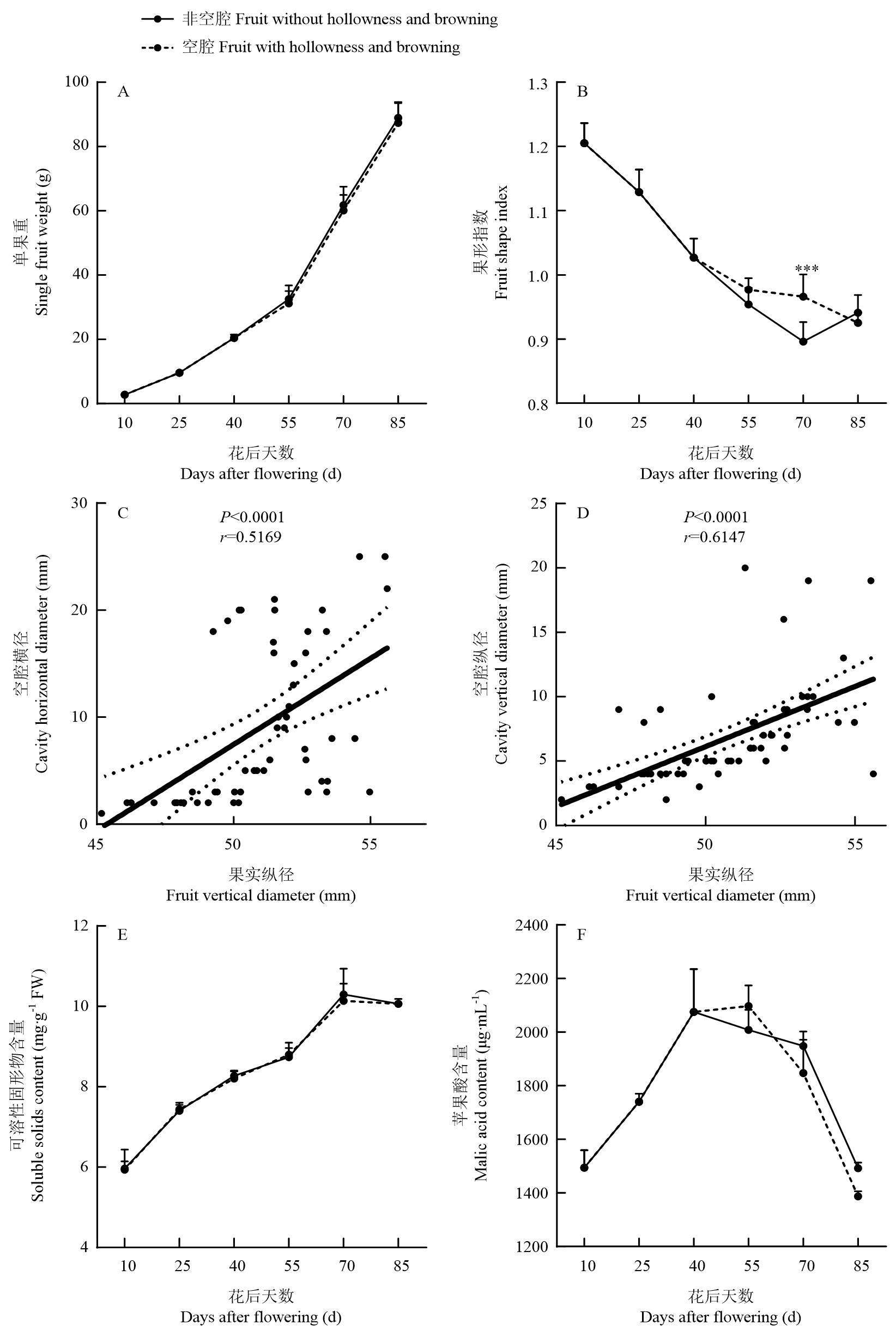
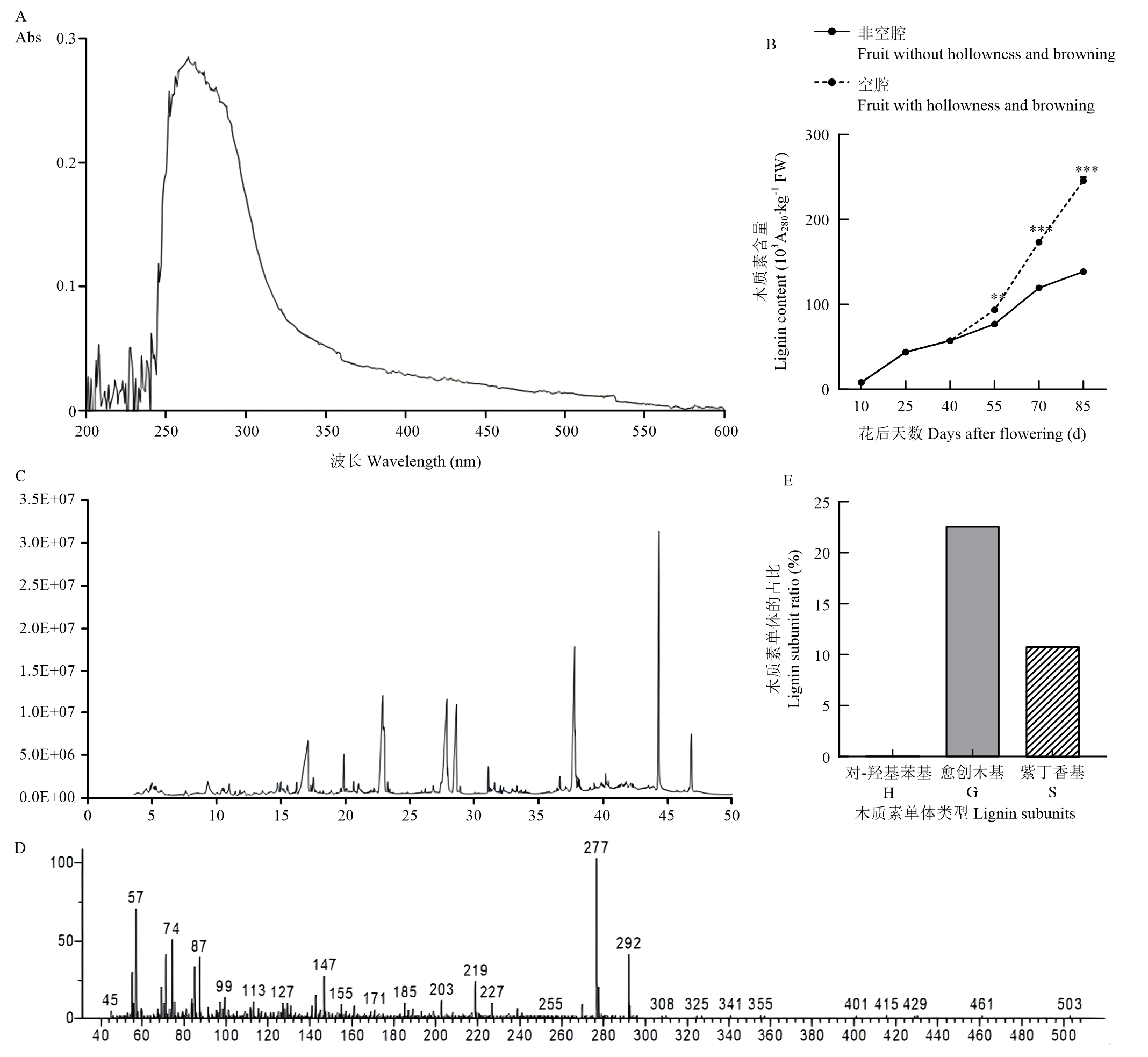
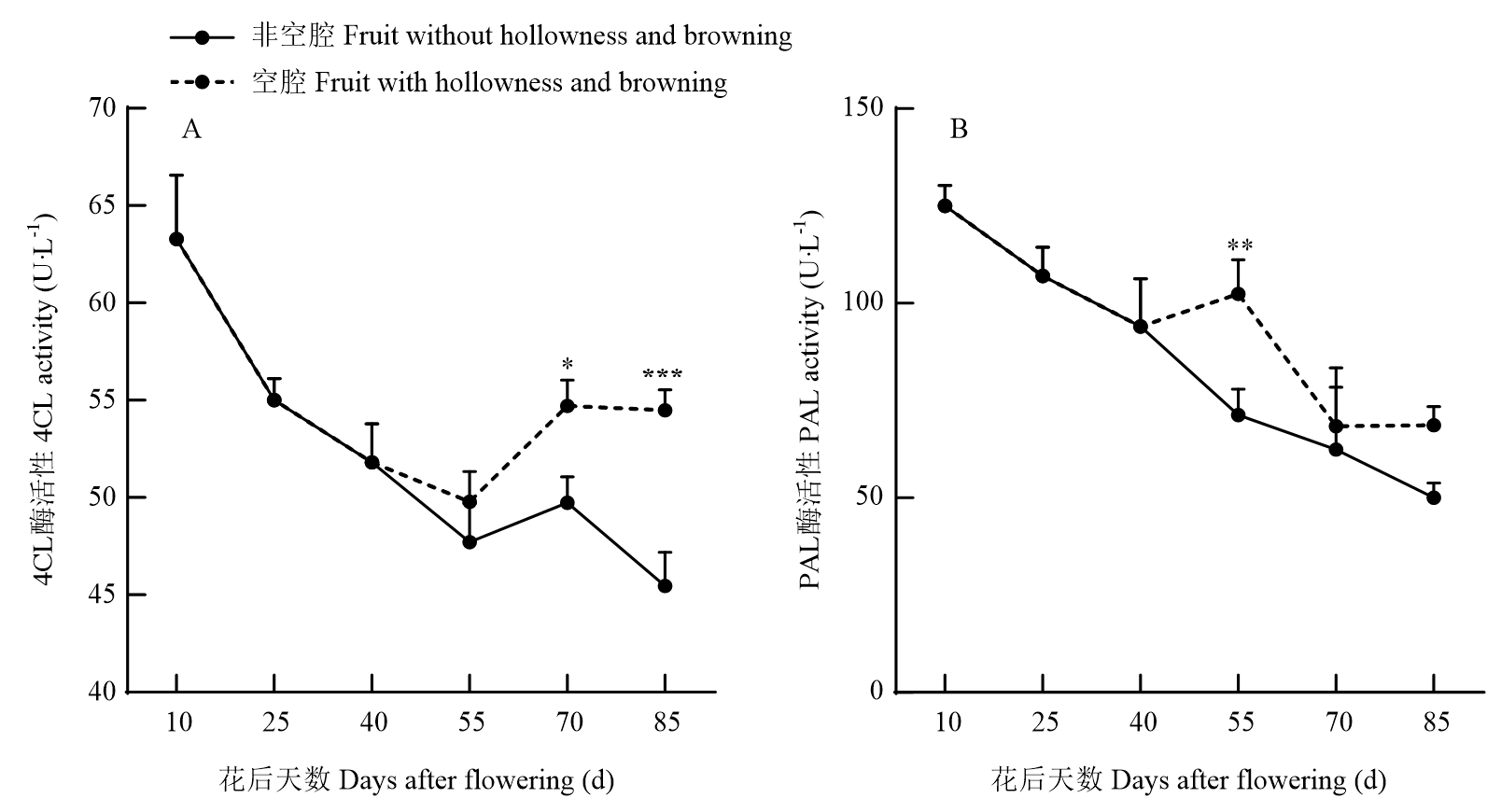
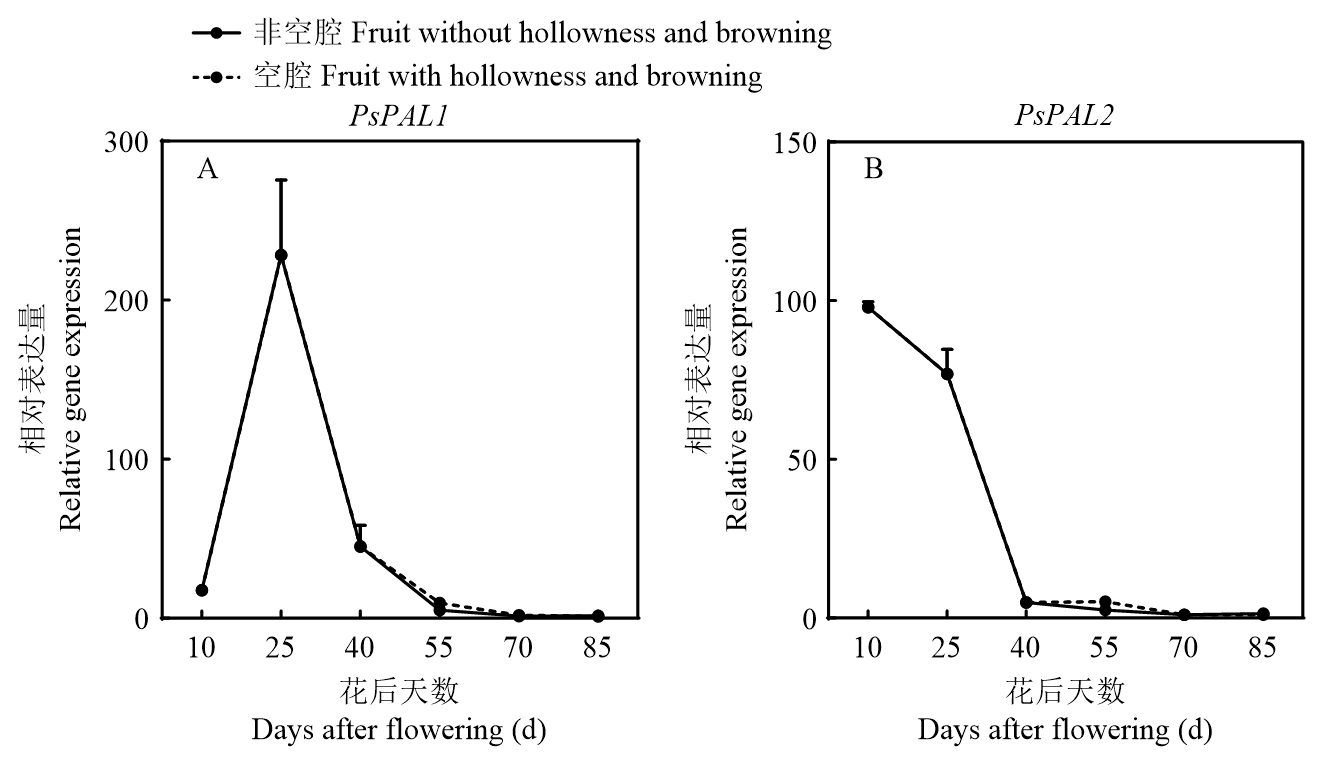
Scientia Agricultura Sinica ›› 2025, Vol. 58 ›› Issue (4): 759-778.doi: 10.3864/j.issn.0578-1752.2025.04.011
• HORTICULTURE • Previous Articles Next Articles
LI JianKun( ), PENG Chao, ZHANG ZiYang, LIANG Xi, WEI MingKang, YANG Qiang, LI BinQi, Muhammad Moaaz Ali, Viola Kayima, CHEN FaXing(), DENG HongHong()
), PENG Chao, ZHANG ZiYang, LIANG Xi, WEI MingKang, YANG Qiang, LI BinQi, Muhammad Moaaz Ali, Viola Kayima, CHEN FaXing(), DENG HongHong()
| [1] |
张加延. 中国果树科学与实践-李. 西安: 陕西科学技术出版社, 2015.
|
|
|
|
| [2] |
刘硕, 徐铭, 刘家成, 章秋平, 马小雪, 刘宁, 张玉萍, 张玉君, 赵海娟, 刘威生. 世界李育种概况. 中国农业科学, 2023, 56(9): 1744-1759. doi: 10.3864/j.issn.0578-1752.2023.09.011.
|
|
|
|
| [3] |
王立如, 范林洁, 房聪玲, 张建春, 沈群超, 吴月燕. 植物生长调节剂对阳光玫瑰葡萄果实品质的影响. 浙江农业科学, 2023, 64(3): 639-645.
doi: 10.16178/j.issn.0528-9017.20220617 |
|
|
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
苍涛, 宋雯, 赵学平, 吴长兴, 吴声敢, 陈丽萍, 徐明飞, 赵慧宇, 王强. 氯吡脲对草莓空心与畸形的影响. 浙江农业学报, 2017, 29(9): 1537-1543.
doi: 10.3969/j.issn.1004-1524.2017.09.16 |
|
doi: 10.3969/j.issn.1004-1524.2017.09.16 |
|
| [11] |
|
| [12] |
赖灯妮, 张群, 尚雪波, 谭欢, 周雨佳, 郭薇丹, 付湘晋. 氯吡脲处理对6种猕猴桃品质和货架期的影响. 食品安全质量检测学报, 2023, 14(1): 146-155.
|
|
|
|
| [13] |
杨旭风, 贾晓东, 许梦洋, 莫正海, 贾展慧, 宣继萍. 褐变机理及其防治技术研究进展. 中国农学通报, 2023, 39(13): 137-145.
doi: 10.11924/j.issn.1000-6850.casb2022-0400 |
|
doi: 10.11924/j.issn.1000-6850.casb2022-0400 |
|
| [14] |
邱栋梁, 刘星辉. 果肉的褐变机理. 福建农业大学学报, 1998, 27(1): 37-41.
|
|
|
|
| [15] |
邱栋梁, 刘星辉. 奈贮藏过程中果肉褐变的研究. 农业工程学报, 1999, 15(1): 253-254.
|
|
|
|
| [16] |
姜翠翠. 㮈多酚氧化酶(PPO)基因启动子的克隆与功能鉴定[D]. 福州: 福建农林大学, 2008.
|
|
|
|
| [17] |
姜翠翠. 㮈均一化全长cDNA文库的构建及果实褐变相关基因的分离与表达分析[D]. 福州: 福建农林大学, 2012.
|
|
|
|
| [18] |
|
| [19] |
李笑月, 袁艳文, 顾晨冉, 秦智伟, 辛明. 不同环境因子和基因型对黄瓜空腔性状的影响. 北方园艺, 2023, 16: 18-25.
|
|
|
|
| [20] |
|
| [21] |
USDA. United States standards for grades of watermelons. (2006) [December 22, 2023]. https://permanent.fdlp.gov/LPS92640/WatermelonStandard.pdf.
|
| [22] |
USDA. United States standards for grades of watermelons (2021) [December 22, 2023] https://www.ams.usda.gov/sites/default/files/media/WatermelonStandards.pdf.
|
| [23] |
doi: S0308-8146(19)30526-6 pmid: 30955619 |
| [24] |
|
| [25] |
薛维文, 周显芳, 张昭其, 方方. 果蔬采后木质素积累及其调控对品质的影响研究进展. 园艺学报, 2022, 49(9): 2023-2036.
doi: 10.16420/j.issn.0513-353x.2021-0268 |
|
doi: 10.16420/j.issn.0513-353x.2021-0268 |
|
| [26] |
doi: S0958-1669(18)30143-5 pmid: 30913460 |
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
doi: 10.1007/s00425-018-2965-z pmid: 30078075 |
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [1] | WANG JiaNuo, CHEN GuiPing, LI Pan, WANG LiPing, NAN YunYou, HE Wei, FAN ZhiLong, HU FaLong, CHAI Qiang, YIN Wen, ZHAO LiaoHao. Photo-Physiological Mechanism at Grain Filling Stage of No-Tillage with Plastic Re-Mulching to Increase Maize Yield in Oasis Irrigation Areas [J]. Scientia Agricultura Sinica, 2026, 59(6): 1189-1202. |
| [2] | LÜ XuDong, SUN ShiYuan, LI YaNan, LIU YuLong, WANG YanQun, FU Xin, ZHANG JiaYing, NING Peng, PENG ZhengPing. Effects of Intelligent Mechanized Layered Fertilization on Root-Soil Nutrient Distribution and Yield in Wheat Fields [J]. Scientia Agricultura Sinica, 2026, 59(1): 129-146. |
| [3] | CHEN GuiPing, LI Pan, SHAO GuanGui, WU XiaYu, YIN Wen, ZHAO LianHao, FAN ZhiLong, HU FaLong. The Regulatory Effect of Reduced Irrigation and Combined Organic- Inorganic Fertilizer Application on Stay-Green Characteristics in Silage Maize Leaves After Tasseling Stage [J]. Scientia Agricultura Sinica, 2025, 58(7): 1381-1396. |
| [4] | WU WenQi, JIAO Yang, XI JiaZhen, WANG XuFeng, GUO BoSen, SHEN YuFang. Effects of Different Organic Materials Combined with Chemical Fertilizer on Soil Fertility and Maize Grain Yield [J]. Scientia Agricultura Sinica, 2025, 58(23): 4966-4978. |
| [5] | WU DongMing, XU Jing, YUAN JiaMei, WANG KeXuan, LI YuYi, HE Ping, ZHANG JianFeng, SONG DaLi, GAO Miao, ZHOU Wei. Isolation and Identification of Rhizosphere Growth-Promoting Bacteria of Myroides odoratimimus PJ-3 and Their Salt/ Alkali-Tolerance and Growth-Promoting Effects on Maize [J]. Scientia Agricultura Sinica, 2025, 58(20): 4131-4143. |
| [6] | ZHANG HaiRui, JIA AngYuan, GAO QiQi, HAN ZheQun, NAN ShanShan, DUAN BiHua, WU XuePing. The Effects of Biochar Combined with Fulvic Acid on the Physical and Chemical Properties, Enzyme Activities and Multifunctionality of Soil in Coastal Saline-Alkali Land [J]. Scientia Agricultura Sinica, 2025, 58(20): 4178-4188. |
| [7] | XUAN ZePeng, FENG HuiYao, CHEN MeiQi, XU JiSheng, LIU MengXuan, ZHAO BingZi, ZHANG JiaBao. Effects of Straw Returning Combined with Chemical Fertilizer on Soil Ecosystem Multifunctionality [J]. Scientia Agricultura Sinica, 2025, 58(14): 2821-2837. |
| [8] | ZHAO YuMeng, LI GuangXu, DAI QiDong, LIU JiaCheng, ZHAO HaiJuan, ZHANG YuJun, YANG Hua, LIU Shuo. Isolation and Identification of Pathogen of Plum Bacterial Shot Hole and Laboratory Test of Bactericide Toxicity [J]. Scientia Agricultura Sinica, 2025, 58(12): 2358-2370. |
| [9] | LI AoGui, CHEN YaRu, PENG ZiYang, RU JingYi, LIU WeiXing. Litter Decomposition Characteristics of Steppe Ecosystems with Different Precipitation Gradients [J]. Scientia Agricultura Sinica, 2025, 58(1): 117-126. |
| [10] | GAO YaFei, ZHAO YuanBo, XU Lin, SUN JiaYue, XIA YuXuan, XUE Dan, WU HaiWen, NING Hang, WU AnChi, WU Lin. Long-Term Sphagnum Cultivation in Cold Waterlogged Paddy Fields Increases Organic Carbon Content and Decreases Soil Extracellular Enzyme Activities [J]. Scientia Agricultura Sinica, 2024, 57(8): 1533-1546. |
| [11] | ZHAO HaiJuan, ZHANG YuPing, ZHANG YuJun, LIU Ning, XU Ming, LIU JiaCheng, WANG BiJun, LIU WeiSheng, LIU Shuo. Evaluation of Fruit Aroma in Chinese Plum Germplasm Based on Electronic Nose Technology [J]. Scientia Agricultura Sinica, 2024, 57(21): 4328-4341. |
| [12] | SHAO JiaZhu, LÜ Wen, LIAO XinLin, YUAN XinYu, SONG Zhen, JIANG DongHua. Isolation and Identification of Soybean Rhizosphere Growth-Promoting Bacteria and Their Salt Tolerance and Growth-Promoting Effects [J]. Scientia Agricultura Sinica, 2024, 57(21): 4248-4263. |
| [13] | LIU Shuo, XU Ming, LIU JiaCheng, ZHANG QiuPing, MA XiaoXue, LIU Ning, ZHANG YuPing, ZHANG YuJun, ZHAO HaiJuan, LIU WeiSheng. An Overview of the Worldwide Plum Breeding [J]. Scientia Agricultura Sinica, 2023, 56(9): 1744-1759. |
| [14] | DONG Xiu, ZHANG Yan, MUNYAMPIRWA Tito, TAO HaiNing, SHEN YuYing. Effects of Long-Term Conservation Tillage on Soil Carbon Content and Invertase Activity in Dry Farmland on the Loess Plateau [J]. Scientia Agricultura Sinica, 2023, 56(5): 907-919. |
| [15] | ZHANG LingFei, MA Lei, LI YuDong, ZHENG FuLi, WEI JianLin, TAN DeShui, CUI XiuMin, LI Yan. Effects of Long-Term Synergistic Application of Organic Materials and Chemical Fertilizers on Bacterial Community and Enzyme Activity in Wheat-Maize Rotation Fluvo-Aquic Soil [J]. Scientia Agricultura Sinica, 2023, 56(19): 3843-3855. |
|
||