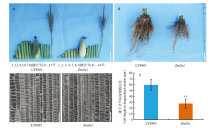
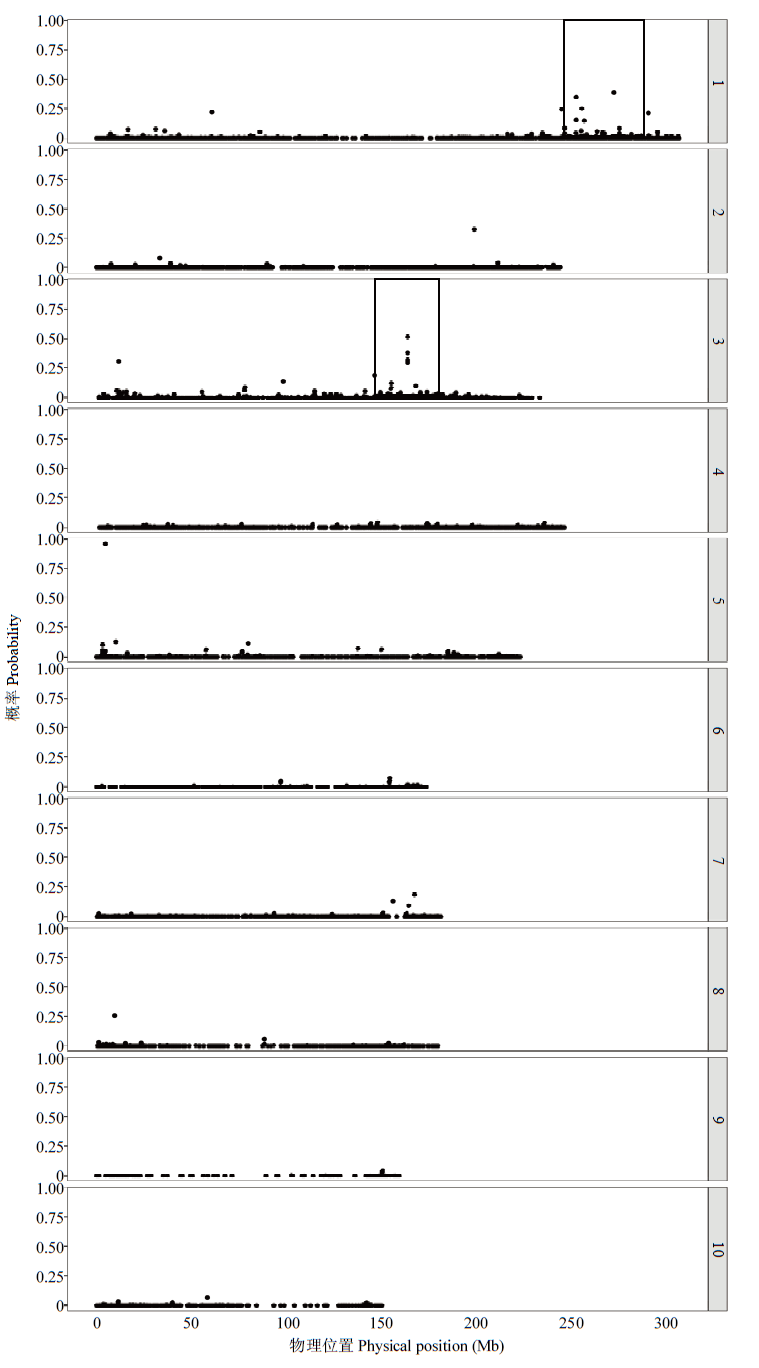
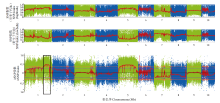
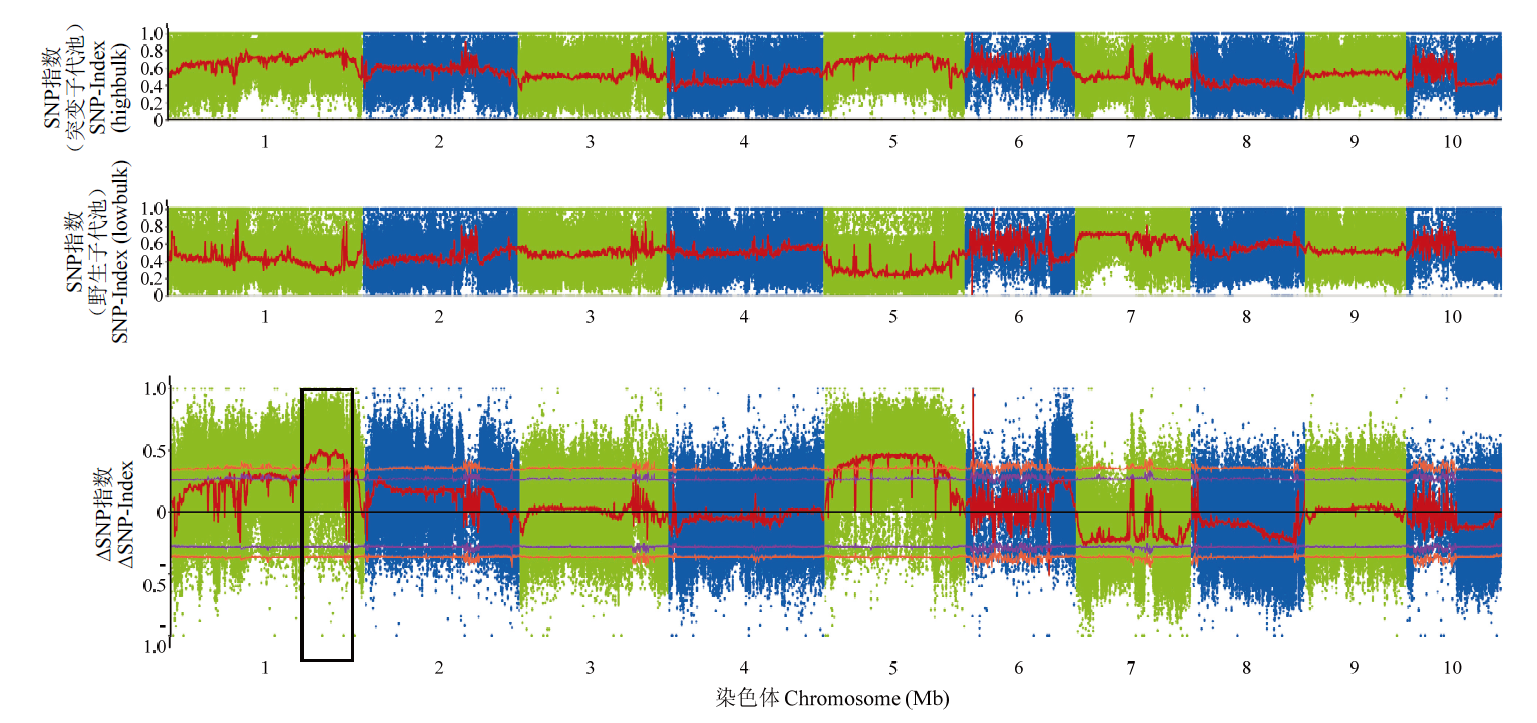
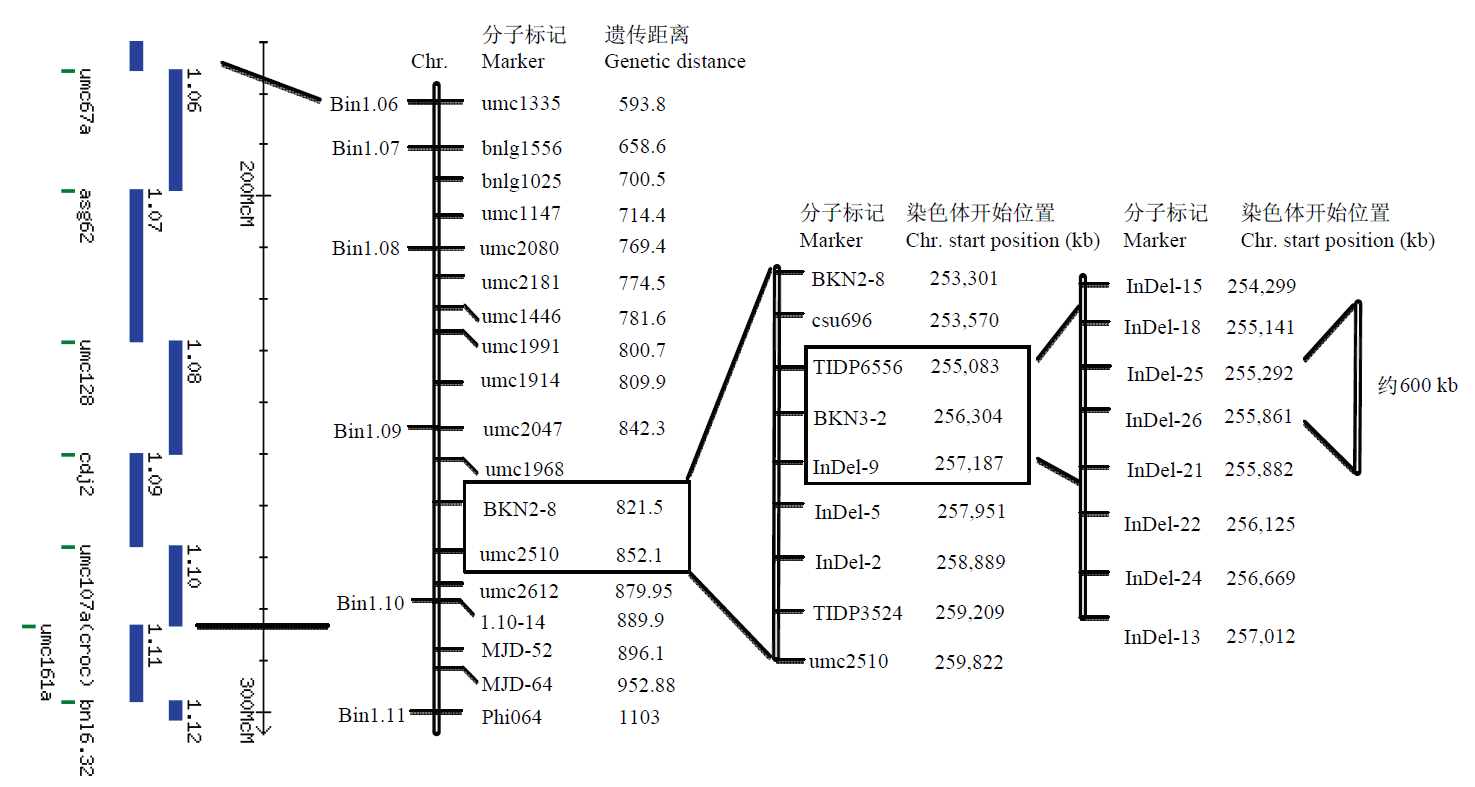
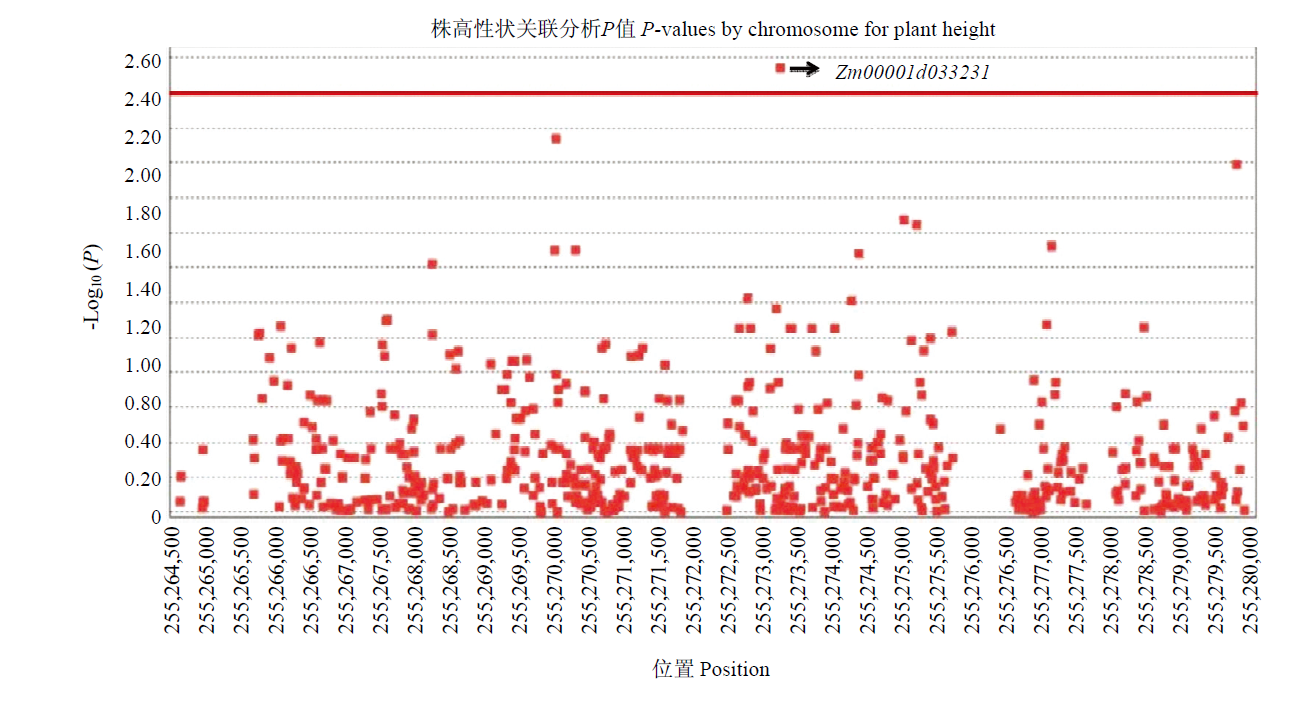
Scientia Agricultura Sinica ›› 2023, Vol. 56 ›› Issue (5): 821-837.doi: 10.3864/j.issn.0578-1752.2023.05.002
• CROP GENETICS & BREEDING · GERMPLASM RESOURCES · MOLECULAR GENETICS • Previous Articles Next Articles
ZHOU WenQi1( ), ZHANG HeTong2, HE HaiJun1, GONG DianMing2, YANG YanZhong1, LIU ZhongXiang1, LI YongSheng1, WANG XiaoJuan1, LIAN XiaoRong1, ZHOU YuQian1(), QIU FaZhan2()
), ZHANG HeTong2, HE HaiJun1, GONG DianMing2, YANG YanZhong1, LIU ZhongXiang1, LI YongSheng1, WANG XiaoJuan1, LIAN XiaoRong1, ZHOU YuQian1(), QIU FaZhan2()
| [1] |
周文期, 周玉乾, 刘忠祥, 连晓荣, 王晓娟, 何海军, 杨彦忠, 寇思荣. 12C6+重离子束辐照玉米后代的生物学效应. 核农学报, 2019, 33(12): 2311-2318.
doi: 10.11869/j.issn.100-8551.2019.12.2311 |
|
|
|
| [2] |
周文期, 王晓娟, 寇思荣, 李艳春, 何海军. 玉米叶夹角形成的分子调控机理研究. 土壤与作物, 2019, 8(3): 339-348.
|
|
|
|
| [3] |
doi: 10.1111/tpj.12038 pmid: 23020630 |
| [4] |
doi: 10.1038/416701a |
| [5] |
|
| [6] |
doi: 10.1038/22307 |
| [7] |
doi: 10.1534/genetics.113.159152 pmid: 24514905 |
| [8] |
doi: 10.1007/s00122-011-1551-6 pmid: 21359559 |
| [9] |
doi: 10.1126/science.1086072 |
| [10] |
doi: 10.1093/jxb/erv182 pmid: 25922491 |
| [11] |
周文期, 连晓荣, 刘忠祥, 周玉乾, 赵小强. 玉米株高和穗位高的调控机理研究. 分子植物育种, 2021, 19(23): 7965-7976.
|
|
|
|
| [12] |
马雅杰, 高悦欣, 李依萍, 龙艳, 董振营, 万向元. 玉米株高和穗位高的遗传基础与分子机制. 中国生物工程杂志, 2021, 41(12): 61-73.
|
|
|
|
| [13] |
doi: 10.1371/journal.pone.0030798 |
| [14] |
doi: 10.1038/nature12583 |
| [15] |
doi: 10.1038/90135 pmid: 11431702 |
| [16] |
doi: 10.1002/ajb2.1955.42.issue-10 |
| [17] |
|
| [18] |
doi: 10.1111/pbi.v18.6 |
| [19] |
doi: 10.1105/tpc.109.068221 |
| [20] |
doi: 10.1016/s0031-9422(02)00656-8 pmid: 12591256 |
| [21] |
doi: 10.1126/science.144.3620.849 |
| [22] |
doi: 10.1104/pp.114.247486 pmid: 25341533 |
| [23] |
doi: 10.1104/pp.112.204503 pmid: 22961131 |
| [24] |
|
| [25] |
doi: 10.1104/pp.88.4.1367 |
| [26] |
doi: 10.1093/jxb/erv554 pmid: 26767748 |
| [27] |
doi: 10.1104/pp.16.00399 |
| [28] |
doi: 10.1242/dev.01671 |
| [29] |
doi: 10.1093/pcp/pcq153 pmid: 20937610 |
| [30] |
doi: 10.1105/tpc.10.11.1875 |
| [31] |
doi: 10.1371/journal.pone.0037040 |
| [32] |
doi: 10.1105/tpc.110.075267 pmid: 21335375 |
| [33] |
|
| [34] |
doi: 10.1104/pp.15.00367 pmid: 26162429 |
| [35] |
|
| [36] |
|
| [37] |
doi: 10.1111/pbi.12751 pmid: 28499064 |
| [38] |
doi: 10.1126/science.273.5280.1406 pmid: 8703079 |
| [39] |
doi: 10.1007/s11103-005-1674-8 |
| [40] |
周文期, 寇思荣, 连晓荣, 杨彦忠, 刘忠祥, 王晓娟, 何海军, 周玉乾. 水稻和玉米叶表皮突变体的筛选和鉴定. 植物生理学报, 2020, 56(2): 189-199.
|
|
doi: 10.1111/ppl.1982.56.issue-2 |
|
| [41] |
周文期, 连晓荣, 周玉乾, 王兴荣, 杨彦忠, 刘忠祥, 王晓娟, 何海军, 寇思荣. EMS诱变玉米自交系种质创新应用. 玉米科学, 2020, 28(6): 31-38.
|
|
|
|
| [42] |
doi: 10.1038/nbt.2095 pmid: 22267009 |
| [43] |
周文期, 刘忠祥, 王晓娟, 何海军, 周玉乾, 杨彦忠, 连晓荣, 李永生. 利用BSA-Seq方法快速定位作物农艺性状QTL/基因概述. 甘肃农业科技, 2022, 53(4): 1-10.
|
|
|
|
| [44] |
doi: 10.1007/s10681-006-9312-3 |
| [45] |
doi: 10.1186/1471-2229-11-4 pmid: 21211047 |
| [46] |
doi: 10.1007/s11103-013-0129-x pmid: 24214124 |
| [47] |
孙强, 任姣姣, 徐晓明, 李宗泽, 黄博文, 陈占辉, 吴鹏昊. 玉米株高和穗位高QTL定位和全基因组选择探究. 玉米科学, 2022, 30(4): 40-47.
|
|
|
|
| [48] |
doi: 10.1007/s11032-009-9298-3 |
| [49] |
doi: 10.1016/s1369-5266(00)00211-9 pmid: 11641069 |
| [50] |
doi: S1369-5266(17)30199-1 pmid: 30142487 |
| [51] |
doi: 10.1074/jbc.L120.013410 |
| [52] |
doi: 10.1105/tpc.011965 |
| [53] |
|
| [54] |
doi: 10.1242/dev.030353 pmid: 19633168 |
| [55] |
doi: 10.1093/mp/ssr034 pmid: 21505044 |
| [56] |
doi: 10.1093/pcp/pcm024 pmid: 17303594 |
| [57] |
doi: 10.1111/pbi.v18.8 |
| [58] |
doi: 10.1111/j.1467-7652.2011.00637.x pmid: 21777365 |
| [59] |
|
| [1] | MA Nan, AN TingTing, ZHANG JiuMing, WANG JingKuan. Effects of Maize Shoot and Root Residues Added on Microbial Residue Carbon and Nitrogen in Different Fertility Levels of Black Soil [J]. Scientia Agricultura Sinica, 2023, 56(4): 686-696. |
| [2] | LIU Dan, AN YuLi, TAO XiaoXiao, WANG XiaoZhong, LÜ DianQiu, GUO YanJun, CHEN XinPing, ZHANG WuShuai. Effects of Different Nitrogen Gradients on Yield and Nitrogen Uptake of Hybrid Seed Maize in Northwest China [J]. Scientia Agricultura Sinica, 2023, 56(3): 441-452. |
| [3] | ZHAO ZhengXin,WANG XiaoYun,TIAN YaJie,WANG Rui,PENG Qing,CAI HuanJie. Effects of Straw Returning and Nitrogen Fertilizer Types on Summer Maize Yield and Soil Ammonia Volatilization Under Future Climate Change [J]. Scientia Agricultura Sinica, 2023, 56(1): 104-117. |
| [4] | CHAI HaiYan,JIA Jiao,BAI Xue,MENG LingMin,ZHANG Wei,JIN Rong,WU HongBin,SU QianFu. Identification of Pathogenic Fusarium spp. Causing Maize Ear Rot and Susceptibility of Some Strains to Fungicides in Jilin Province [J]. Scientia Agricultura Sinica, 2023, 56(1): 64-78. |
| [5] | LI ZhouShuai,DONG Yuan,LI Ting,FENG ZhiQian,DUAN YingXin,YANG MingXian,XU ShuTu,ZHANG XingHua,XUE JiQuan. Genome-Wide Association Analysis of Yield and Combining Ability Based on Maize Hybrid Population [J]. Scientia Agricultura Sinica, 2022, 55(9): 1695-1709. |
| [6] | XIONG WeiYi,XU KaiWei,LIU MingPeng,XIAO Hua,PEI LiZhen,PENG DanDan,CHEN YuanXue. Effects of Different Nitrogen Application Levels on Photosynthetic Characteristics, Nitrogen Use Efficiency and Yield of Spring Maize in Sichuan Province [J]. Scientia Agricultura Sinica, 2022, 55(9): 1735-1748. |
| [7] | LI YiLing,PENG XiHong,CHEN Ping,DU Qing,REN JunBo,YANG XueLi,LEI Lu,YONG TaiWen,YANG WenYu. Effects of Reducing Nitrogen Application on Leaf Stay-Green, Photosynthetic Characteristics and System Yield in Maize-Soybean Relay Strip Intercropping [J]. Scientia Agricultura Sinica, 2022, 55(9): 1749-1762. |
| [8] | MA XiaoYan,YANG Yu,HUANG DongLin,WANG ZhaoHui,GAO YaJun,LI YongGang,LÜ Hui. Annual Nutrients Balance and Economic Return Analysis of Wheat with Fertilizers Reduction and Different Rotations [J]. Scientia Agricultura Sinica, 2022, 55(8): 1589-1603. |
| [9] | LI Qian,QIN YuBo,YIN CaiXia,KONG LiLi,WANG Meng,HOU YunPeng,SUN Bo,ZHAO YinKai,XU Chen,LIU ZhiQuan. Effect of Drip Fertigation Mode on Maize Yield, Nutrient Uptake and Economic Benefit [J]. Scientia Agricultura Sinica, 2022, 55(8): 1604-1616. |
| [10] | LIU Shuo,ZHANG Hui,GAO ZhiYuan,XU JiLi,TIAN Hui. Genetic Variations of Potassium Harvest Index in 437 Wheat Varieties [J]. Scientia Agricultura Sinica, 2022, 55(7): 1284-1300. |
| [11] | ZHANG JiaHua,YANG HengShan,ZHANG YuQin,LI CongFeng,ZHANG RuiFu,TAI JiCheng,ZHOU YangChen. Effects of Different Drip Irrigation Modes on Starch Accumulation and Activities of Starch Synthesis-Related Enzyme of Spring Maize Grain in Northeast China [J]. Scientia Agricultura Sinica, 2022, 55(7): 1332-1345. |
| [12] | ZHI Lei,ZHE Li,SUN NanNan,YANG Yang,Dauren Serikbay,JIA HanZhong,HU YinGang,CHEN Liang. Genome-Wide Association Analysis of Lead Tolerance in Wheat at Seedling Stage [J]. Scientia Agricultura Sinica, 2022, 55(6): 1064-1081. |
| [13] | TAN XianMing,ZHANG JiaWei,WANG ZhongLin,CHEN JunXu,YANG Feng,YANG WenYu. Prediction of Maize Yield in Relay Strip Intercropping Under Different Water and Nitrogen Conditions Based on PLS [J]. Scientia Agricultura Sinica, 2022, 55(6): 1127-1138. |
| [14] | QIAO Yuan,YANG Huan,LUO JinLin,WANG SiXian,LIANG LanYue,CHEN XinPing,ZHANG WuShuai. Inputs and Ecological Environment Risks Assessment of Maize Production in Northwest China [J]. Scientia Agricultura Sinica, 2022, 55(5): 962-976. |
| [15] | LIU Miao,LIU PengZhao,SHI ZuJiao,WANG XiaoLi,WANG Rui,LI Jun. Critical Nitrogen Dilution Curve and Nitrogen Nutrition Diagnosis of Summer Maize Under Different Nitrogen and Phosphorus Application Rates [J]. Scientia Agricultura Sinica, 2022, 55(5): 932-947. |
|
||