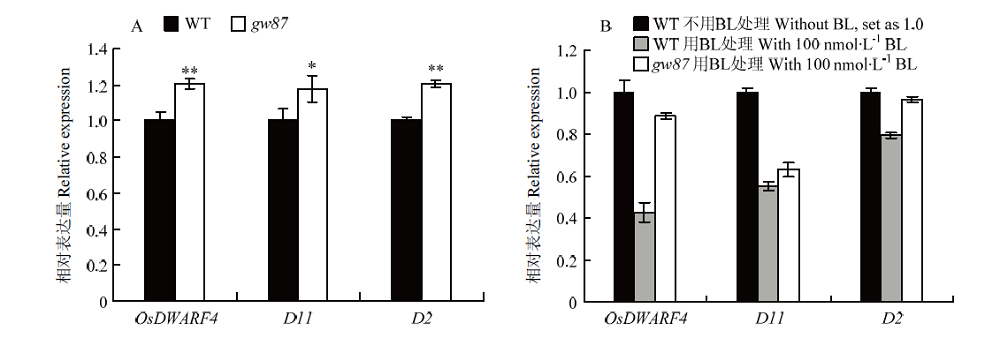
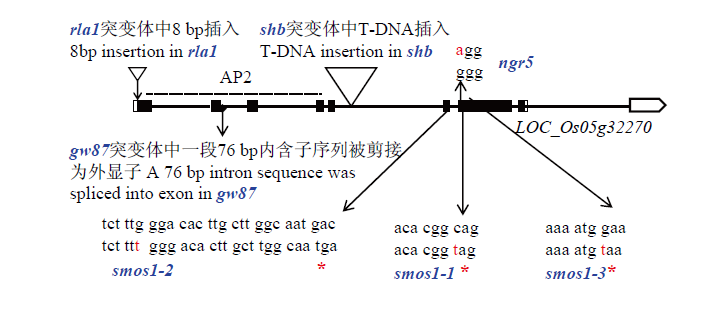
Scientia Agricultura Sinica ›› 2021, Vol. 54 ›› Issue (12): 2487-2498.doi: 10.3864/j.issn.0578-1752.2021.12.001
• CROP GENETICS & BREEDING·GERMPLASM RESOURCES·MOLECULAR GENETICS • Previous Articles Next Articles
ZHANG XiangYu( ),GUO Jia,WANG San,CHEN CongPing,SUN ChangHui,DENG XiaoJian,WANG PingRong()
),GUO Jia,WANG San,CHEN CongPing,SUN ChangHui,DENG XiaoJian,WANG PingRong()
| [1] |
TANKSLEY S D. Mapping polygenes. Annual Review of Genetics, 1993,27(1):205-233.
doi: 10.1146/annurev.ge.27.120193.001225 |
| [2] |
YANO M. Genetic and molecular dissection of naturally occurring variation. Current Opinion in Plant Biology, 2001,4(2):130-135.
doi: 10.1016/S1369-5266(00)00148-5 |
| [3] |
SONG X J, HUANG W, SHI M, ZHU M Z, LIN H X. A QTL for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase. Nature Genetics, 2007,39(5):623-630.
doi: 10.1038/ng2014 |
| [4] |
SHOMURA A, IZAWA T, EBANA K, EBITANI T, KANEGAE H, KONISHI S, YANO M. Deletion in a gene associated with grain size increased yields during rice domestication. Nature Genetics, 2008,40(8):1023-1028.
doi: 10.1038/ng.169 |
| [5] |
WENG J F, GU S H, WAN X Y, GAO H, GUO T, SU N, LEI C L, ZHANG X, CHENG Z J, GUO X P, WANG J L, JIANG L, ZHAI H Q, WAN J M. Isolation and initial characterization of GW5, a major QTL associated with rice grain width and weight. Cell Research, 2008,18(12):1199-1209.
doi: 10.1038/cr.2008.307 |
| [6] |
LI Y B, FAN C C, XING Y Z, JIANG Y H, LUO L J, SUN L, SHAO D, XU C J, LI X H, XIAO J H, HE Y Q, ZHANG Q F. Natural variation in GS5 plays an important role in regulating grain size and yield in rice. Nature Genetics, 2011,43(12):1266-1269.
doi: 10.1038/ng.977 |
| [7] |
XU C J, LIU Y, LI Y B, XU X D, XU C G, LI X H, XIAO J H, ZHANG Q F. Differential expression of GS5 regulates grain size in rice. Journal of Experimental Botany, 2015,66(9):2611-2623.
doi: 10.1093/jxb/erv058 |
| [8] |
WANG S K, WU K, YUAN Q B, LIU X Y, LIU Z B, LIN X Y, ZENG R Z, ZHU H T, DONG G J, QIAN Q, ZHANG G Q, FU X D. Control of grain size, shape and quality by OsSPL16 in rice. Nature Genetics, 2012,44(8):950-954.
doi: 10.1038/ng.2327 |
| [9] |
WANG S K, LI S, LIU Q, WU K, ZHANG J Q, WANG S S, WANG Y, CHEN X B, ZHANG Y, GAO C X, WANG F, HUANG H X, FU X D. The OsSPL16-GW7 regulatory module determines grain shape and simultaneously improves rice yield and grain quality. Nature Genetics, 2015,47(8):949-954.
doi: 10.1038/ng.3352 |
| [10] |
TONG H N, CHU C C. Functional specificities of brassinosteroid and potential utilization for crop improvement. Trends in Plant Science, 2018,23(11):1016-1028.
doi: 10.1016/j.tplants.2018.08.007 |
| [11] | PLANAS-RIVEROLA A, GUPTA A, BETEGÓN-PUTZE I, BOSCH N, IBAÑES M, CAÑO-DELGADO A I. Brassinosteroid signaling in plant development and adaptation to stress. Development,2019,146, dev151894. |
| [12] |
SAKAMOTO T, MORINAKA Y, OHNISHI T, SUNOHARA H, FUJIOKA S, UEGUCHI-TANAKA M, MIZUTANI M, SAKATA K, TAKATSUTO S, YOSHIDA S, TANAKA H, KITANO H, MATSUOKA M. Erect leaves caused by brassinosteroid deficiency increase biomass production and grain yield in rice. Nature Biotechnology, 2006,24(1):105-109.
doi: 10.1038/nbt1173 |
| [13] |
HONG Z, UEGUCHI-TANAKA M, UMEMURA K, UOZU S, FUJIOKA S, TAKATSUTO S, YOSHIDA S, ASHIKARI M, KITANO H, MATSUOKA M. A rice brassinosteroid-deficient mutant, ebisu dwarf (d2), is caused by a loss of function of a new member of cytochrome P450. The Plant Cell, 2003,15(12):2900-2910.
doi: 10.1105/tpc.014712 |
| [14] |
TANABE S, ASHIKARI M, FUJIOKA S, TAKATSUTO S, YOSHIDA S, YANO M, YOSHIMURA A, KITANO H, MATSUOKA M, FUJISAWA Y, KATO H, IWASAKI Y. A novel cytochrome P450 is implicated in brassinosteroid biosynthesis via the characterization of a rice dwarf mutant, dwarf11, with reduced seed length. The Plant Cell, 2005,17(3):776-790.
doi: 10.1105/tpc.104.024950 |
| [15] |
HONG Z, UEGUCHI-TANAKA M, SHIMIZU-SATO S, INUKAI Y, FUJIOKA S, SHIMADA Y, TAKATSUTO S, AGETSUMA M, YOSHIDA S, WATANABE Y, UOZU S, KITANO H, ASHIKARI M, MATSUOKA M. Loss-of-function of a rice brassinosteroid biosynthetic enzyme, C-6 oxidase, prevents the organized arrangement and polar elongation of cells in the leaves and stem. The Plant Journal, 2002,32(4):495-508.
doi: 10.1046/j.1365-313X.2002.01438.x |
| [16] |
LIU X, FENG Z M, ZHOU C L, REN Y K, MOU C L, WU T, YANG C Y, LIU S J, JIANG L, WAN J M. Brassinosteroid (BR) biosynthetic gene lhdd10 controls late heading and plant height in rice (Oryza sativa L.). Plant Cell Reports, 2016,35(2):357-368.
doi: 10.1007/s00299-015-1889-3 |
| [17] |
YAMAMURO C, IHARA Y, WU X, NOGUCHI T, FUJIOKA S, TAKATSUTO S, ASHIKARI M, KITANO H, MATSUOKA M. Loss of function of a rice brassinosteroid insensitive1 homolog prevents internode elongation and bending of the lamina joint. The Plant Cell, 2000,12(9):1591-1605.
doi: 10.1105/tpc.12.9.1591 |
| [18] |
MORINAKA Y, SAKAMOTO T, INUKAI Y, AGETSUMA M, KITANO H, ASHIKARI M, MATSUOKA M. Morphological alteration caused by brassinosteroid insensitivity increases the biomass and grain production of rice. Plant Physiology, 2006,141:924-931.
doi: 10.1104/pp.106.077081 |
| [19] |
CHE R H, TONG H N, SHI B H, LIU Y Q, FANG S R, LIU D P, XIAO Y H, HU B, LIU L C, WANG H R, ZHAO M F, CHU C C. Control of grain size and rice yield by GL2-mediated brassinosteroid responses. Nature Plants, 2015,2(1):15195.
doi: 10.1038/nplants.2015.195 |
| [20] |
TONG H N, LIU L C, JIN Y, DU L, YIN Y H, QIAN Q, ZHU L H, CHU C C. DWARF AND LOW-TILLERING acts as a direct downstream target of a GSK3/SHAGGY-like kinase to mediate brassinosteroid responses in rice. The Plant Cell, 2012,24(6):2562-2577.
doi: 10.1105/tpc.112.097394 |
| [21] |
SUN L J, LI X J, FU Y C, ZHU Z F, TAN L B, LIU F X, SUN X Y, SUN X W, SUN C Q. GS6, a member of the gras gene family, negatively regulates grain size in rice. Journal of Integrative Plant Biology, 2013,55(10):938-949.
doi: 10.1111/jipb.12062 |
| [22] |
QIAO S L, SUN S Y, WANG L L, WU Z H, LI C X, LI X M, WANG T, LENG L N, TIAN W S, LU T G, WANG X L. The RLA1/SMOS1 transcription factor functions with OsBZR1 to regulate brassinosteroid signaling and rice architecture. The Plant Cell, 2017,29(2):292-309.
doi: 10.1105/tpc.16.00611 |
| [23] |
AYA K, HOBO T, SATO-IZAWA K, UEGUCHI-TANAKA M, KITANO H, MATSUOKA M. A novel AP2-type transcription factor, SMALL ORGAN SIZE1, controls organ size downstream of an auxin signaling pathway. Plant and Cell Physiology, 2014,55(5):897-912.
doi: 10.1093/pcp/pcu023 |
| [24] |
HIRANO K, YOSHIDA H, AYA K, KAWAMURA M, HAYASHI M, HOBO T, SATO-IZAWA K, KITANO H, UEGUCHI-TANAKA M, MATSUOKA M. SMALL ORGAN SIZE 1 and SMALL ORGAN SIZE 2/DWARF AND LOW-TILLERING form a complex to integrate auxin and brassinosteroid signaling in rice. Molecular Plant, 2017,10(4):590-604.
doi: 10.1016/j.molp.2016.12.013 |
| [25] | TONG H N, CHU C C. Physiological analysis of brassinosteroid responses and sensitivity in rice. Methods in Molecular Biology, 2017,1564:23-29. |
| [26] |
LICHTENTHALER H K, WELLBURN A R. Determination of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochemical Society Transactions, 1983,11(5):591-592.
doi: 10.1042/bst0110591 |
| [27] | MCCOUCH S R, KOCHERT G, YU Z H, WANG Z Y, KHUSH G S, COFFMANW R, TANKLEY S D. Molecular mapping of rice chromosomes. The Oretical and Applied Genet, 1988,76(6):815-829. |
| [28] |
LI J T, ZHAO Y, CHU H W, WANG L K, FU Y R, LIU P, UPADHYAYA N, CHEN C L, MOU T M, FENG Y Q, KUMAR P, XU J. SHOEBOX modulates root meristem size in rice through dose-dependent effects of gibberellins on cell elongation and proliferation. PLoS Genetics, 2015,11(8):e1005464.
doi: 10.1371/journal.pgen.1005464 |
| [29] | WU K, WANG S S, SONG W Z, ZHANG J Q, WANG Y, LIU Q, YU J P, YE Y F, LI S, CHEN J F, ZHAO Y, WANG J, WU X K, WANG M Y, ZHANG Y J, LIU B M, WU Y J, HARBERD N P, FU X D. Enhanced sustainable green revolution yield via nitrogen-responsive chromatin modulation in rice. Science, 2020,367(6478): eaaz2046. |
| [1] | WANG JiaNuo, CHEN GuiPing, LI Pan, WANG LiPing, NAN YunYou, HE Wei, FAN ZhiLong, HU FaLong, CHAI Qiang, YIN Wen, ZHAO LiaoHao. Photo-Physiological Mechanism at Grain Filling Stage of No-Tillage with Plastic Re-Mulching to Increase Maize Yield in Oasis Irrigation Areas [J]. Scientia Agricultura Sinica, 2026, 59(6): 1189-1202. |
| [2] | ZHOU XinJie, REN Hao, CHEN YingLong, ZHANG JiWang, ZHAO Bin, REN BaiZhao, LIU Peng, WANG HongZhang. Effects of Calcium Peroxide on Root Morphology and Yield Formation of Summer Maize in Waterlogging Farmland [J]. Scientia Agricultura Sinica, 2026, 59(6): 1203-1216. |
| [3] | LI SiYuan, LI HongPing, CHANG HongQing, ZHANG SenYan, LI SiJia, CUI XinFei, QIAO Po, ZENG Bo, LIU GuiZhen, LIU TianXue, TANG JiHua, LI ChaoHai. Effects of Density Increase on Dynamic Change of Yield and Agronomic Traits of Maize Cultivars with Different Plant Heights [J]. Scientia Agricultura Sinica, 2026, 59(5): 967-984. |
| [4] | WANG YongSheng, NIU Li, WANG ChangJie, MA LiHua, LIAN XiaoXiao, MENG YaXiong, MA XiaoLe, YAO LiRong, ZHANG Hong, YANG Ke, LI BaoChun, WANG HuaJun, SI ErJing, WANG JunCheng. Genome-Wide Association Study and Candidate Gene Identification for Thousand Grain Weight in Winter Wheat [J]. Scientia Agricultura Sinica, 2026, 59(3): 499-514. |
| [5] | LI XinYi, LI JiaNing, YANG WenPing, XIA Qing, HUO YingRui, HAO ShiHang, HUANG TingMiao, REN YongKang, CHEN Jie, GAO ZhiQiang, YANG ZhenPing. Effects of Post-Anthesis Foliar Zinc Application on Zinc Nutrition in Colored-Grain Wheat [J]. Scientia Agricultura Sinica, 2026, 59(3): 515-527. |
| [6] | LÜ XuDong, SUN ShiYuan, LI YaNan, LIU YuLong, WANG YanQun, FU Xin, ZHANG JiaYing, NING Peng, PENG ZhengPing. Effects of Intelligent Mechanized Layered Fertilization on Root-Soil Nutrient Distribution and Yield in Wheat Fields [J]. Scientia Agricultura Sinica, 2026, 59(1): 129-146. |
| [7] | FEI YaoYing, WANG Di, TANG WeiJie, GUO CaiLi, ZHANG XiaoHu, QIU XiaoLei, CHENG Tao, YAO Xia, JIANG ChongYa, ZHU Yan, CAO WeiXing, ZHENG HengBiao. Estimation of Rice Grain Protein Content Using Fusion Imagery from UAV-based Multi-Sensors [J]. Scientia Agricultura Sinica, 2026, 59(1): 41-56. |
| [8] | PU LiXia, ZHANG JiaRui, YE JianPing, HUANG XiuLan, FAN GaoQiong, YANG HongKun. The Combined Effects of 16, 17-Dihydro Gibberellin A5 and Straw Mulching on Tillering and Grain Yield of Dryland Wheat [J]. Scientia Agricultura Sinica, 2025, 58(9): 1735-1748. |
| [9] | WEI Ping, PAN JuZhong, ZHU DePing, SHAO ShengXue, CHEN ShanShan, WEI YaQian, GAO WeiWei. The Function of OsDREB1J in Regulating Rice Grain Size [J]. Scientia Agricultura Sinica, 2025, 58(8): 1463-1478. |
| [10] | LIU JinSong, WU LongMei, BAO XiaoZhe, LIU ZhiXia, ZHANG Bin, YANG TaoTao. Effects of a Short-Term Reduction in Nitrogen Fertilizer Application Rates on the Grain Yield and Rice Quality of Early and Late-Season Dual-Use Rice in South China [J]. Scientia Agricultura Sinica, 2025, 58(8): 1508-1520. |
| [11] | XUE YuQi, ZHAO JiYu, SUN WangSheng, REN BaiZhao, ZHAO Bin, LIU Peng, ZHANG JiWang. Effects of Different Nitrogen Forms on Yield and Quality of Summer Maize [J]. Scientia Agricultura Sinica, 2025, 58(8): 1535-1549. |
| [12] | YIN Bo, YU AiZhong, WANG PengFei, YANG XueHui, WANG YuLong, SHANG YongPan, ZHANG DongLing, LIU YaLong, LI Yue, WANG Feng. Effects of Green Manure Returning Combined with Nitrogen Fertilizer Reduction on Hydrothermal Characteristics of Wheat Field and Grain Yield in Oasis Irrigation Area [J]. Scientia Agricultura Sinica, 2025, 58(7): 1366-1380. |
| [13] | SHE WenTing, SUN RuiQing, DANG HaiYan, LI WenHu, ZHANG Feng, TIAN Yi, XU JunFeng, DING YuLan, WANG ZhaoHui. Sulfur Concentration and Distribution in Wheat Grain Sampled from Farmers’ Fields in Main Wheat Production Regions of China and Its Affecting Factors [J]. Scientia Agricultura Sinica, 2025, 58(5): 956-974. |
| [14] | LI Lu, XIE Zhuang, XIE KeYing, ZHANG Han, ZHAO ZhuoWen, XIANG AoNi, LI QiaoLong, LING YingHua, HE GuangHua, ZHAO FangMing. Construction of Single and Dual-Segment Substitution Lines from Rice CSSL-Z492 and Genetic Dissection of QTL for Grain Size [J]. Scientia Agricultura Sinica, 2025, 58(3): 401-415. |
| [15] | QIU HaiLong, LI Pan, ZHANG DianKai, FAN ZhiLong, HU FaLong, CHEN GuiPing, FAN Hong, HE Wei, YIN Wen, ZHAO LianHao. Compensatory Effects of Multiple Cropping Green Manure on Growth and Yield Loss of Nitrogen-Reduced Spring Wheat in Oasis Irrigation Areas of Northwest China [J]. Scientia Agricultura Sinica, 2025, 58(3): 443-459. |
|
||