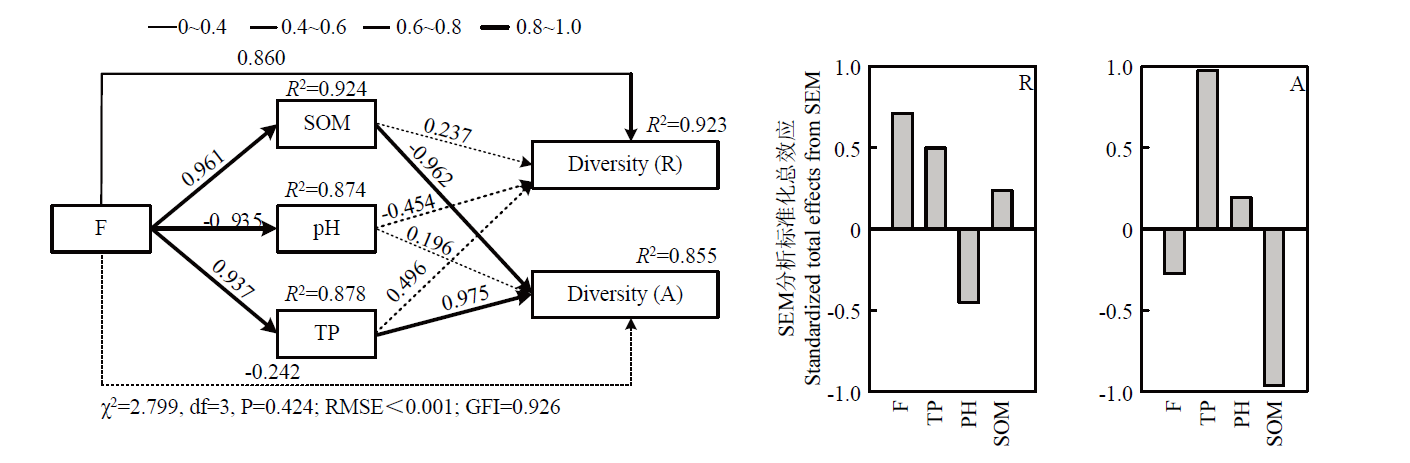
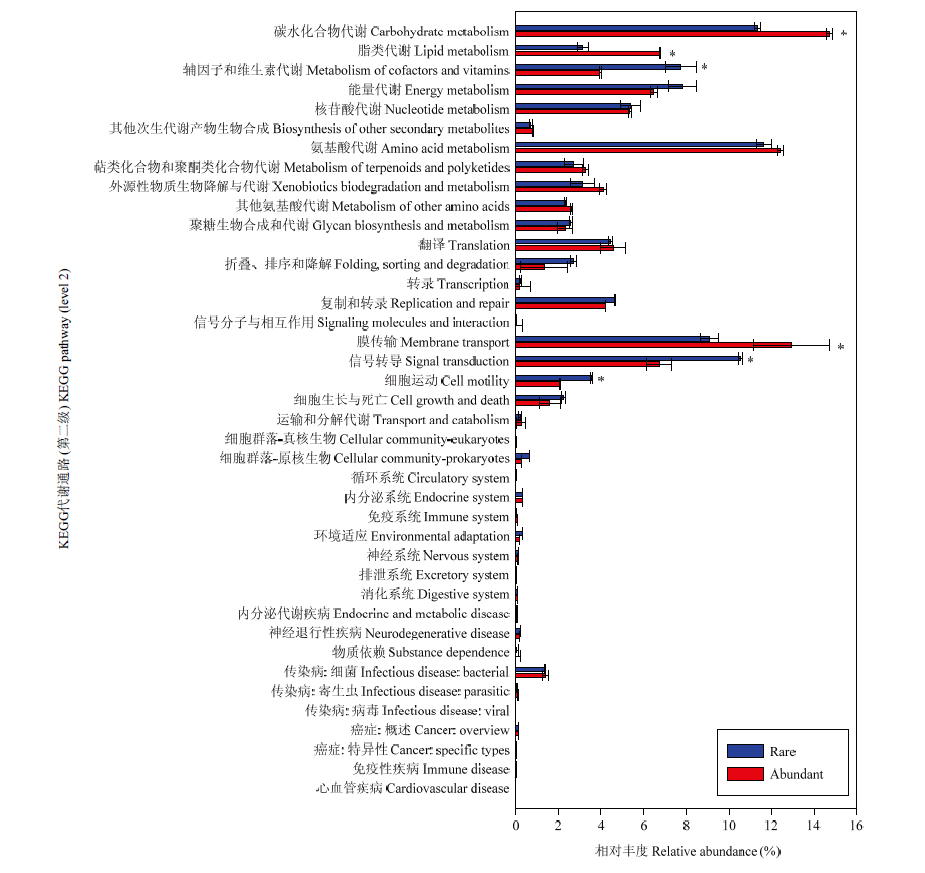
Scientia Agricultura Sinica ›› 2023, Vol. 56 ›› Issue (18): 3615-3628.doi: 10.3864/j.issn.0578-1752.2023.18.010
• SOIL & FERTILIZER·WATER-SAVING IRRIGATION·AGROECOLOGY & ENVIRONMENT • Previous Articles Next Articles
LIU Lei( ), SHI JianShuo(), ZHANG GuoYin, GAO Jing, LI Pin, REN Yanli, WANG LiYing()
), SHI JianShuo(), ZHANG GuoYin, GAO Jing, LI Pin, REN Yanli, WANG LiYing()
| [1] |
doi: 10.1126/science.1153213 |
| [2] |
doi: S0966-842X(18)30047-7 pmid: 29550356 |
| [3] |
doi: 10.1038/s41396-018-0153-6 |
| [4] |
王敬国. 设施菜田退化土壤修复与资源高效利用. 北京: 中国农业大学出版社, 2011.
|
|
|
|
| [5] |
doi: 10.1016/j.apsoil.2017.08.016 |
| [6] |
doi: 10.1007/s11356-017-9837-0 |
| [7] |
doi: 10.1007/s00248-021-01741-8 |
| [8] |
doi: 10.3389/fmicb.2018.02096 pmid: 30283408 |
| [9] |
doi: 10.1038/s41396-018-0159-0 |
| [10] |
doi: 10.1038/ismej.2015.29 |
| [11] |
doi: 10.1016/j.agee.2018.11.025 |
| [12] |
doi: 10.1007/s00374-019-01355-w |
| [13] |
doi: 10.1111/1462-2920.14945 pmid: 32067386 |
| [14] |
doi: 10.1111/mec.14218 pmid: 28665016 |
| [15] |
doi: 10.1038/nrmicro3400 pmid: 25730701 |
| [16] |
|
| [17] |
|
| [18] |
doi: S0269-7491(16)32803-2 pmid: 28336094 |
| [19] |
doi: 10.1007/s11356-021-16830-x |
| [20] |
邢肖毅, 倪绯, 张亚丽, 黎颖惠, 杨贤均, 尹丹红. 增温对土壤丰富和稀有微生物的差异性影响. 环境科学与技术, 2022, 45(5): 70-76.
|
|
doi: 10.1021/es101444k |
|
| [21] |
doi: 10.1016/j.cub.2014.02.050 |
| [22] |
doi: 10.1126/science.1182570 pmid: 20150447 |
| [23] |
doi: 10.1016/j.soilbio.2019.107686 |
| [24] |
姜灿烂, 何园球, 刘晓利, 陈平帮, 王艳玲, 李辉信. 长期施用有机肥对旱地红壤团聚体结构与稳定性的影响. 土壤学报, 2010, 47(4): 715-722.
|
|
|
|
| [25] |
鲍士旦. 土壤农化分析. 3版. 北京: 中国农业出版社, 2000.
|
|
|
|
| [26] |
doi: 10.1016/j.soilbio.2018.08.016 |
| [27] |
doi: 10.1016/j.soilbio.2021.108222 |
| [28] |
doi: 10.3390/d11060098 |
| [29] |
doi: 10.1016/j.tim.2006.04.007 |
| [30] |
|
| [31] |
关健飞, 沈智超, 曹阳. 稀有微生物群落研究进展. 湖北农业科学, 2020, 59(15): 5-11.
|
|
|
|
| [32] |
doi: 10.1111/emi.v23.8 |
| [33] |
doi: 10.1111/1462-2920.13098 pmid: 26486414 |
| [34] |
doi: S0045-6535(19)31415-8 pmid: 31260865 |
| [35] |
doi: 10.1186/s40168-020-00892-z |
| [36] |
doi: 10.1038/s41587-019-0328-3 pmid: 31792408 |
| [37] |
pmid: 17479464 |
| [1] | WANG Feng, CHANG YunNi, WU ZhiDan, SUN Jun, JIANG FuYing, CHEN YuZhen, YU WenQuan. Effects of Long-Term Nitrogen Application on Soil Fungal Diversity, Functional Groups and Assembly Processes in Tea Gardens [J]. Scientia Agricultura Sinica, 2026, 59(2): 368-385. |
| [2] | HE ZhiLin, SUN CuiXia, YUE HongLi, TAN YueXia, ZHANG YaoHai, WANG FuSheng, LIU SiTao, JIANG Dong. Genetic Diversity Analysis and GWAS of Alloocimene Based on Resequencing of Citron, Lemon Germplasm Resources [J]. Scientia Agricultura Sinica, 2026, 59(2): 386-401. |
| [3] | BAI YuXin, LIU LingZhi, AN TingTing, LI ShuangYi, WANG JingKuan. Eeffects of Long-Term Fertilization on Bacterial Community Structure and Carbon Metabolic Functions in Brown Soil [J]. Scientia Agricultura Sinica, 2025, 58(8): 1579-1590. |
| [4] | SHI Fan, LI WenGuang, YI ShuSheng, YANG Na, CHEN YuMeng, ZHENG Wei, ZHANG XueChen, LI ZiYan, ZHAI BingNian. The Variation Characteristics of Soil Organic Carbon Fractions Under the Combined Application of Organic and Inorganic Fertilizers [J]. Scientia Agricultura Sinica, 2025, 58(4): 719-732. |
| [5] | XU YuJuan, ZHANG Jie, WANG TianYi, CHEN HaoYang, ZHAO JiaJia, WU BangBang, HAO YuQiong, LI XiaoHua, ZHENG XingWei, ZUO JingJing, ZHENG Jun. Identification of Glu-A3 and Glu-B3 of Low-Molecular-Weight Glutenin in Shanxi Wheat and Its Effect on Quality [J]. Scientia Agricultura Sinica, 2025, 58(24): 5110-5127. |
| [6] | CHEN JunQuan, MA ChiYuan, HU Xin, LI Duo, GUO YanQi, LIU Can, ZHOU Kai, ZHENG TaiHui. Effects of Incorporation of Inorganic-Organic Fertilizers on Soil Fertility, Ecological Stoichiometric Characteristics, and Yields of Rice Cropping System in the Red Soil Region of China [J]. Scientia Agricultura Sinica, 2025, 58(23): 4952-4966. |
| [7] | CHEN CaiJin, MA Lin, JIANG QingXue, LIU JinHui, MIAO Tong, ZHANG ZhiPeng, MENG Xiang, MA XiaoRan, ZHOU XinYue, ZHANG Jian, LIU WenHui, WANG XueMin. Genetic Diversity Analysis of Phenotypic Traits of 244 Forage Oat Germplasm Resources [J]. Scientia Agricultura Sinica, 2025, 58(23): 4825-4836. |
| [8] | WEI YiMin, ZHOU MeiLiang, TANG Yu. Origin, Evolution and Spread of Crop Buckwheat [J]. Scientia Agricultura Sinica, 2025, 58(21): 4305-4316. |
| [9] | LIU XiaoXu, ZHONG ZeXin, QIU JiaRen, YANG ChunXiao, ZHANG YongJun, XIE Wen, ZHANG YouJun, PAN HuiPeng. GENETIC DIVERSITY OF MTCO1 IN DIFFERENT GEOGRAPHICAL POPULATIONS OF MEGALUROTHRIPS USITATUS [J]. Scientia Agricultura Sinica, 2025, 58(21): 4361-4371. |
| [10] | XIE HaiPeng, LIN JunXu, LIU Yong, MAI XianJun, LUO Feng, WANG XueWu, XIE Wen, LI ShaoKa, KONG XiangYi, WU XiaoYan. Effects of Different Organic Fertilizers on the Control of Cowpea Wilt by Bacillus velezensis SD13 [J]. Scientia Agricultura Sinica, 2025, 58(21): 4405-4420. |
| [11] | TANG GuiMei, LI WeiDong, ZHOU YuXia, KONG YouHan, XIAO XiaoLing, PENG YingShu, ZHANG Li, FU HongYan, LIU Yang, HUANG GuoLin. Genetic Diversity Analysis of Cymbidium faberi Germplasm Resources Based on Phenotypic Traits [J]. Scientia Agricultura Sinica, 2025, 58(2): 339-354. |
| [12] | LI YueQi, MA ZhongHua, SU Ming, LIU Hao, MA FengLan, MA XiaoYing, LI Tao, LI QingYun, ZHANG Dan, LIU JiLi, WU Na. Response of Maize Photosynthetic Production Capacity in Saline- Alkaline Soil to Organic Fertilizer Application Rates Under Differential Tillage Practices [J]. Scientia Agricultura Sinica, 2025, 58(19): 3872-3889. |
| [13] | GUO MengZe, ZHANG Lei, SUN PingPing, JIANG Biao, YAN JinQiang, LI ZhengNan. Molecular Characterization and Evolutionary Dynamics of Tomato Leaf Curl New Delhi Virus Isolate from Wax Gourd (Benincasa hispida) in Guangdong [J]. Scientia Agricultura Sinica, 2025, 58(19): 3890-3904. |
| [14] | LI XueFeng, WANG Hui, ZHANG NingBo, JIN TaiHua, ZHANG ShuEr, ZHENG QuanSheng, TAO JiaShu, LI QingKe, LÜ ShenJin, LI YongZhu. Prediction and Analysis of Feeding Density on Production Performance, Cecal Flora Diversity, Short-Chain Fatty Acid Content and Microbial Differential Function of Langya Chickens [J]. Scientia Agricultura Sinica, 2025, 58(17): 3544-3560. |
| [15] | FANG YaTing, ZHAO Jian, SHENG QianNan, LI KaiXu, WANG XiangHua, ZHANG YangYang, ZHU Jun, CONG RiHuan, LU ZhiFeng, LI XiaoKun, REN Tao, LU JianWei. Effects of Long-Term Chemical Fertilizer and Organic Material Application on Crop Yield and Nutrient Utilization in Rice-Rapeseed Rotation System [J]. Scientia Agricultura Sinica, 2025, 58(16): 3164-3177. |
|
||