Scientia Agricultura Sinica ›› 2023, Vol. 56 ›› Issue (16): 3051-3061.doi: 10.3864/j.issn.0578-1752.2023.16.001
• CROP GENETICS & BREEDING·GERMPLASM RESOURCES·MOLECULAR GENETICS • Previous Articles Next Articles
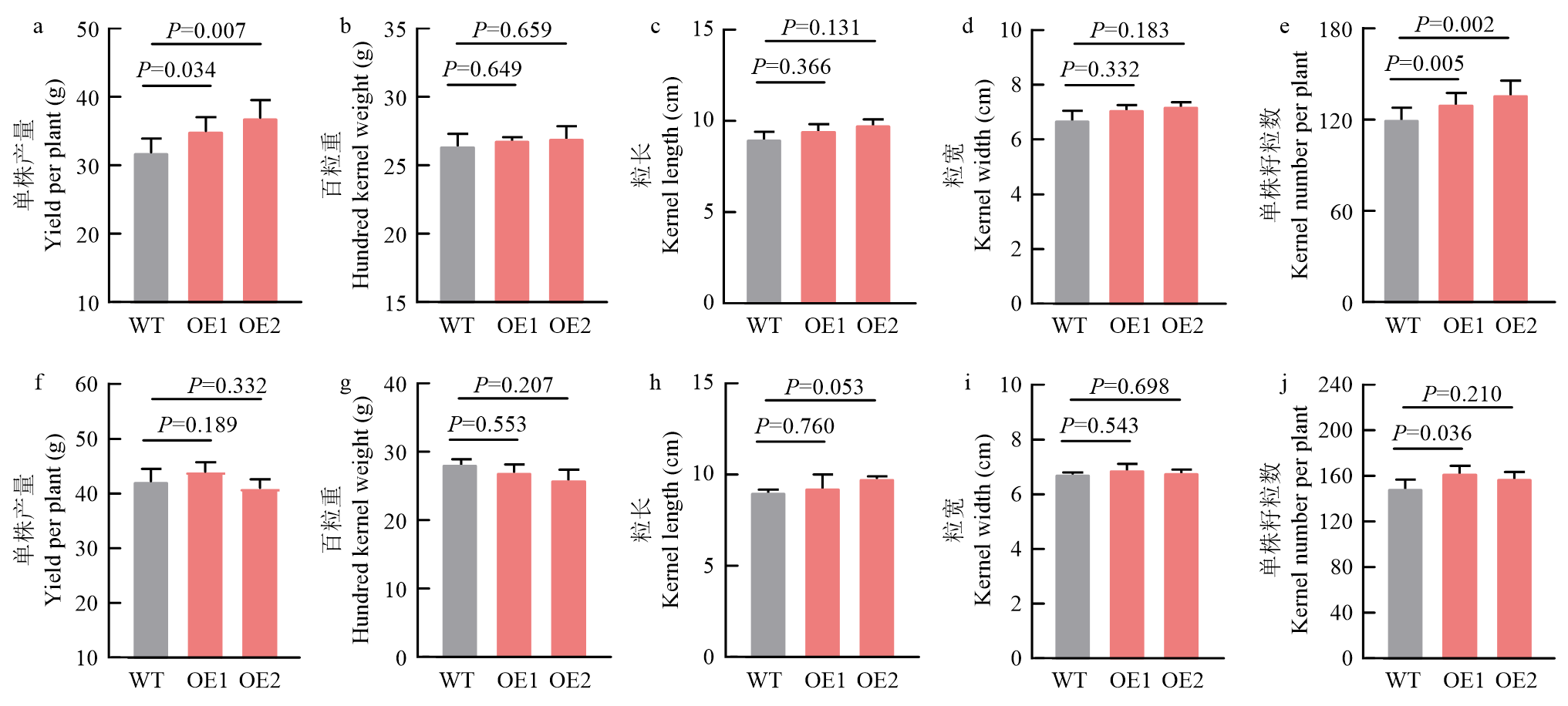
LI Yan1( ), TAO KeYu2(), HU Yue3, LI YongXiang3, ZHANG DengFeng3, LI ChunHui3, HE GuanHua3, SONG YanChun3, SHI YunSu3, LI Yu3, WANG TianYu3, ZOU HuaWen1(), LIU XuYang3()
), TAO KeYu2(), HU Yue3, LI YongXiang3, ZHANG DengFeng3, LI ChunHui3, HE GuanHua3, SONG YanChun3, SHI YunSu3, LI Yu3, WANG TianYu3, ZOU HuaWen1(), LIU XuYang3()
| [1] |
戴景瑞, 鄂立柱. 我国玉米育种科技创新问题的几点思考. 玉米科学, 2010, 18(1): 1-5.
|
|
|
|
| [2] |
|
| [3] |
doi: 10.1126/science.286.5446.1962 |
| [4] |
doi: 10.1126/science.286.5446.1960 pmid: 10583960 |
| [5] |
doi: 10.3389/fpls.2019.00097 pmid: 30815003 |
| [6] |
doi: 10.1016/j.molp.2015.01.007 |
| [7] |
doi: 10.1016/j.cub.2007.05.008 |
| [8] |
doi: 10.1126/science.1141753 pmid: 17446351 |
| [9] |
doi: 10.1038/nature10272 |
| [10] |
doi: 10.1104/pp.107.109538 |
| [11] |
doi: 10.1105/tpc.110.081406 |
| [12] |
doi: 10.1093/jxb/err129 |
| [13] |
doi: 10.1016/j.cub.2018.07.029 |
| [14] |
doi: 10.1104/pp.15.00535 pmid: 26084920 |
| [15] |
doi: S1674-2052(20)30187-8 pmid: 32585190 |
| [16] |
doi: 10.1038/ng.3636 |
| [17] |
doi: 10.1038/s41588-023-01297-y |
| [18] |
doi: 10.1016/j.molp.2016.10.003 |
| [19] |
doi: 10.1038/ncomms9326 |
| [20] |
doi: 10.1093/jxb/ert370 |
| [21] |
doi: 10.1111/pbi.2017.15.issue-2 |
| [22] |
doi: 10.1104/pp.15.00780 |
| [23] |
doi: 10.1093/jxb/eraa507 |
| [24] |
doi: 10.1093/plcell/koab083 |
| [25] |
|
| [26] |
doi: 10.1093/jxb/erw317 |
| [27] |
doi: 10.1111/nph.2017.215.issue-4 |
| [28] |
doi: 10.1080/09168451.2019.1660612 |
| [29] |
doi: 10.1104/pp.19.00676 pmid: 31527087 |
| [30] |
doi: 10.1126/science.abh2890 |
| [1] | MU YingTong, LU JingShi, ZHANG YuTong, SHI FengLing. Identification of Key Drought-Responsive Genes in Upright Medicago ruthenica Sojak cv. Zhilixing Based on Transcriptome Sequencing and WGCNA [J]. Scientia Agricultura Sinica, 2025, 58(21): 4528-4543. |
| [2] | CHEN CaiJin, MA Lin, BAO MingFang, ZHANG GuoHui, JIANG QingXue, YANG TianHui, WANG Chuan, WANG XiaoChun, GAO Ting, WANG XueMin, LIU WenHui. Identification and Evaluation of Drought Resistance for 111 Germplasm Resources of Alfalfa During Germination Stage [J]. Scientia Agricultura Sinica, 2025, 58(10): 1896-1907. |
| [3] | ZHANG YuZhou, WANG YiZhao, GAO RuXi, LIU YiFan. Research Progress on Root System Architecture and Drought Resistance in Wheat [J]. Scientia Agricultura Sinica, 2024, 57(9): 1633-1645. |
| [4] | ZHOU Quan, LU QiuMei, ZHAO ZhangChen, WU ChenRan, FU XiaoGe, ZHAO YuJiao, HAN Yong, LIN HuaiLong, CHEN WeiLin, MOU LiMing, LI XingMao, WANG ChangHai, HU YinGang, CHEN Liang. Identification of Drought Resistance of 244 Spring Wheat Varieties at Seedling Stage [J]. Scientia Agricultura Sinica, 2024, 57(9): 1646-1657. |
| [5] | YAN Wen, JIN XiuLiang, LI Long, XU ZiHan, SU Yue, ZHANG YueQiang, JING RuiLian, MAO XinGuo, SUN DaiZhen. Drought Resistance Evaluation of Synthetic Wheat at Grain Filling Using UAV-Based Multi-Source Imagery Data [J]. Scientia Agricultura Sinica, 2024, 57(9): 1674-1686. |
| [6] | NI ShuHui, SHI DongMei, PAN LiDong, YE Qing, WU JunHao. Response of Cultivated-Layer Water-Holding and Drought Resistance Performance and Productivity to Erosion Degree in Purple Soil Sloping Farmland [J]. Scientia Agricultura Sinica, 2024, 57(7): 1350-1362. |
| [7] | GAO ChenXi, HAO LuYang, HU Yue, LI YongXiang, ZHANG DengFeng, LI ChunHui, SONG YanChun, SHI YunSu, WANG TianYu, LI Yu, LIU XuYang. Analysis of Transposable Element Associated Epigenetic Regulation under Drought in Maize [J]. Scientia Agricultura Sinica, 2024, 57(6): 1034-1048. |
| [8] | ZHU JunJie, ZHANG XinYue, PAN MengYing, ZHANG JingWen, ZHENG Qi, LI YuLing, DONG YongBin. An EIN3/EIL Family Gene, ZmEIL9 Regulates Grain Development in Maize [J]. Scientia Agricultura Sinica, 2024, 57(18): 3522-3532. |
| [9] | HU XueJie, LIU LuPing, WANG FengMin, HAN YuHua, SUN BinCheng, MA QiBin, HUANG ZhiPing, FENG Yan, CHEN Qiang, YANG ChunYan, ZHANG MengChen, ZHANG Kai, QIN Jun. Construction of ms1 Basic Recurrent Populations Adapted to Different Ecological Regions Using Maturity Genes E1 and E2 in Soybean [J]. Scientia Agricultura Sinica, 2024, 57(17): 3305-3317. |
| [10] | CAO LiRu, YE FeiYu, KU LiXia, MA ChenChen, PANG YunYun, LIANG XiaoHan, ZHANG Xin, LU XiaoMin. Analysis of Maize Phenylalanine Ammonia-Lyase (PAL) Family Genes and Functional Study of ZmPAL5 [J]. Scientia Agricultura Sinica, 2024, 57(12): 2265-2281. |
| [11] | LI ShengYou, WANG ChangLing, YAN ChunJuan, ZHANG LiJun, SUN XuGang, CAO YongQiang, WANG WenBin, SONG ShuHong. Evaluation of Drought Resistance in Soybean Germplasm and Identification of Candidate Drought-Resistant Genes [J]. Scientia Agricultura Sinica, 2024, 57(10): 1857-1869. |
| [12] | LUO ZhengYing, HU Xin, LIU XinLong, WU CaiWen, WU ZhuanDi, LIU JiaYong, ZENG QianChun. Application of Trehalose Enhances Drought Resistance in Sugarcane Seedlings and Promotes Plant Growth [J]. Scientia Agricultura Sinica, 2023, 56(21): 4208-4218. |
| [13] | MA YaMei,ZHANG ShaoHong,ZHAO JunLiang,LIU Bin. Function of FCS-Like Zinc-Finger Protein OsFLZ18 in Regulating Rice Flowering Time [J]. Scientia Agricultura Sinica, 2022, 55(20): 3875-3884. |
| [14] | CHANG LiGuo,HE KunHui,LIU JianChao. Mining of Genetic Locus of Maize Stay-Green Related Traits Under Multi-Environments [J]. Scientia Agricultura Sinica, 2022, 55(16): 3071-3081. |
| [15] | MENG Yu,WEN PengFei,DING ZhiQiang,TIAN WenZhong,ZHANG XuePin,HE Li,DUAN JianZhao,LIU WanDai,FENG Wei. Identification and Evaluation of Drought Resistance of Wheat Varieties Based on Thermal Infrared Image [J]. Scientia Agricultura Sinica, 2022, 55(13): 2538-2551. |
|
||