Scientia Agricultura Sinica ›› 2021, Vol. 54 ›› Issue (21): 4500-4513.doi: 10.3864/j.issn.0578-1752.2021.21.002
• CROP GENETICS & BREEDING·GERMPLASM RESOURCES·MOLECULAR GENETICS • Previous Articles Next Articles
ZHU FangFang( ),DONG YaHui(),REN ZhenZhen,WANG ZhiYong,SU HuiHui,KU LiXia,CHEN YanHui()
),DONG YaHui(),REN ZhenZhen,WANG ZhiYong,SU HuiHui,KU LiXia,CHEN YanHui()
| [1] | QU D Y. Food and Agriculture Organization of the United Nations. Viale delle Terme di Caracalla, Rome, Italy. 2016 [June 2021], http://www.fao.org/statistics/databases/en/ . |
| [2] | 代宇佳, 罗晓峰, 周文冠, 陈锋, 帅海威, 杨文钰, 舒凯. 生物和非生物逆境胁迫下的植物系统信号. 植物学报, 2019, 54(2):102-111. |
| DAI Y J, LUO X F, ZHOU W G, CHEN F, SHUAI H W, YANG W Y, SHU K. Plant systemic signaling under biotic and abiotic stresses conditions. Chinese Bulletin of Botany, 2019, 54(2):102-111. (in Chinese) | |
| [3] |
BARTELS D, SUNKAR R. Drought and salt tolerance in plants. Critical Reviews in Plant Sciences, 2005, 24(1):23-58.
doi: 10.1080/07352680590910410 |
| [4] |
MAO H, WANG H, LIU S, LI Z, YANG X, YAN J, LI J, TRAN L, QIN F. A transposable element in a NAC gene is associated with drought tolerance in maize seedlings. Nature Communications, 2015, 6:8326.
doi: 10.1038/ncomms9326 |
| [5] | MAO H, YU L, HAN R, LI Z, LIU H. ZmNAC55, a maize stress-responsive NAC transcription factor, confers drought resistance in transgenic Arabidopsis. Plant Physiology & Biochemistry, 2016, 105:55-66. |
| [6] |
WANG C T, RU J N, LIU Y W, LI M, ZHAO D, YANG J F, FU J D, XU Z S. Maize WRKY transcription factor ZmWRKY106 confers drought and heat tolerance in transgenic plants. International Journal of Molecular Sciences, 2018, 19(10):3046.
doi: 10.3390/ijms19103046 |
| [7] |
WANG C T, RU J N, LIU Y W, YANG J F, MENG L, XU Z S, FU J D. The maize WRKY transcription factor ZmWRKY40 confers drought resistance in transgenic Arabidopsis. International Journal of Molecular Sciences, 2018, 19(9):2580.
doi: 10.3390/ijms19092580 |
| [8] |
YING S, ZHANG D F, JING F, SHI Y S, SONG Y C, WANG T Y, YU L. Cloning and characterization of a maize bZIP transcription factor, ZmbZIP72, confers drought and salt tolerance in transgenic Arabidopsis. Planta, 2012, 235(2):253-266.
doi: 10.1007/s00425-011-1496-7 |
| [9] | 刘彦丹, 英生, 张登峰, 石云素, 宋燕春, 白志川, 王天宇, 黎裕. 玉米逆境胁迫响应基因ZmbZIP71的克隆与表达分析. 植物遗传资源学报, 2011, 12(5):775-781. |
| LIU Y D, YING S, ZHANG D F, SHI Y S, SONG Y C, BAI Z C, WANG T Y, LI Y. Isolation and expression analysis of a stress-responsive gene ZmbZIP71 in maize (Zea mays L.). Journal of Plant Genetic Resources, 2011, 12(5):775-781. (in Chinese) | |
| [10] |
WU J, JIANG Y, LIANG Y, CHEN L, CHEN W, CHENG B. Expression of the maize MYB transcription factor ZmMYB3R enhances drought and salt stress tolerance in transgenic plants. Plant Physiology and Biochemistry, 2019, 137:179-188.
doi: 10.1016/j.plaphy.2019.02.010 |
| [11] |
ZHANG H, XIANG Y, HE N, LIU X, DAI M. Enhanced vitamin C production mediated by an ABA-induced PTP-like nucleotidase improves drought tolerance of Arabidopsis and maize. Molecular Plant, 2020, 13(5):760-776.
doi: 10.1016/j.molp.2020.02.005 |
| [12] |
ZHANG X, MI Y, MAO H, LIU S, QIN F. Genetic variation in ZmTIP1 contributes to root hair elongation and drought tolerance in maize. Plant Biotechnology Journal, 2020, 18(5):1271-1283.
doi: 10.1111/pbi.v18.5 |
| [13] |
DING S, HE F, TANG W, DU H, WANG H. Identification of maize CC-type glutaredoxins that are associated with response to drought stress. Genes, 2019, 10(8):610.
doi: 10.3390/genes10080610 |
| [14] |
LI L, DU Y, HE C, DIETRICH C R, ZHENG J. Maize glossy6 is involved in cuticular wax deposition and drought tolerance. Journal of Experimental Botany, 2019, 70(12):3089-3099.
doi: 10.1093/jxb/erz131 |
| [15] | ZHOU L, ZHOU J, XIONG Y, LIU C, WANG J, WANG G, CAI Y, WU K. Overexpression of a maize plasma membrane intrinsic protein ZmPIP1;1 confers drought and salt tolerance in Arabidopsis. PLoS ONE, 2018, 13(6):e198639. |
| [16] |
WANG H, WANG M, XIA Z. The maize class-I SUMO conjugating enzyme ZmSCE1d is involved in drought stress response. International Journal of Molecular Sciences, 2019, 21(1):29.
doi: 10.3390/ijms21010029 |
| [17] |
LIANG Y, JIANG Y, DU M, LI B, WU J. ZmASR3 from the maize ASR gene family positively regulates drought tolerance in transgenic Arabidopsis. International Journal of Molecular Sciences, 2019, 20(9):2278.
doi: 10.3390/ijms20092278 |
| [18] | FELLER A, MACHEMER K, BRAUN E L, GROTEWOLD E. Evolutionary and comparative analysis of MYB and bHLH plant transcription factors. Plant Journal for Cell & Molecular Biology, 2011, 66(1):94-116. |
| [19] |
WANG F, ZHU H, KONG W, PENG R, LIU Q, YAO Q. The antirrhinum AmDEL gene enhances flavonoids accumulation and salt and drought tolerance in transgenic Arabidopsis. Planta, 2016, 244(1):59-73.
doi: 10.1007/s00425-016-2489-3 |
| [20] | WANG F, ZHU H, CHEN D, LI Z, PENG R, YAO Q. A grape bHLH transcription factor gene, VvbHLH1, increases the accumulation of flavonoids and enhances salt and drought tolerance in transgenic Arabidopsis thaliana. Plant Cell Tissue & Organ Culture, 2016, 125(2):387-398. |
| [21] |
DONG Y, WANG C, HAN X, TANG S, LIU S, XIA X, YIN W. A novel bHLH transcription factor PebHLH35 from Populus euphratica confers drought tolerance through regulating stomatal development, photosynthesis and growth in Arabidopsis. Biochemical and Biophysical Research Communications, 2014, 450(1):453-458.
doi: 10.1016/j.bbrc.2014.05.139 |
| [22] |
CUI X, WANG Y X, LIU Z W, WANG W L, LI H, ZHUANG J. Transcriptome-wide identification and expression profile analysis of the bHLH family genes in Camellia sinensis. Functional and Integrative Genomics, 2018, 18(5):489-503.
doi: 10.1007/s10142-018-0608-x |
| [23] |
LIU W, TAI H, LI S, GAO W, ZHAO M, XIE C, LI W X. bHLH122is important for drought and osmotic stress resistance in Arabidopsis and in the repression of ABA catabolism. New Phytologist, 2014, 201(4):1192-1204.
doi: 10.1111/nph.2014.201.issue-4 |
| [24] | SEO J S, JOO J, KIM M J, KIM Y K, NAHM B H, SANG I S, CHEONG J J, LEE J S, KIM J K, YANG D C. OsbHLH148, a basic helix-loop-helix protein, interacts with OsJAZ proteins in a jasmonate signaling pathway leading to drought tolerance in rice. Plant Journal for Cell & Molecular Biology, 2011, 65(6):907-921. |
| [25] |
LI Z, LIU C, ZHANG Y, WANG B, RAN Q, ZHANG J. The bHLH family member ZmPTF1 regulates drought tolerance in maize by promoting root development and ABA synthesis. Journal of Experimental Botany, 2019, 70(19):5471-5486.
doi: 10.1093/jxb/erz307 |
| [26] |
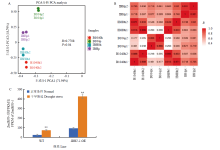
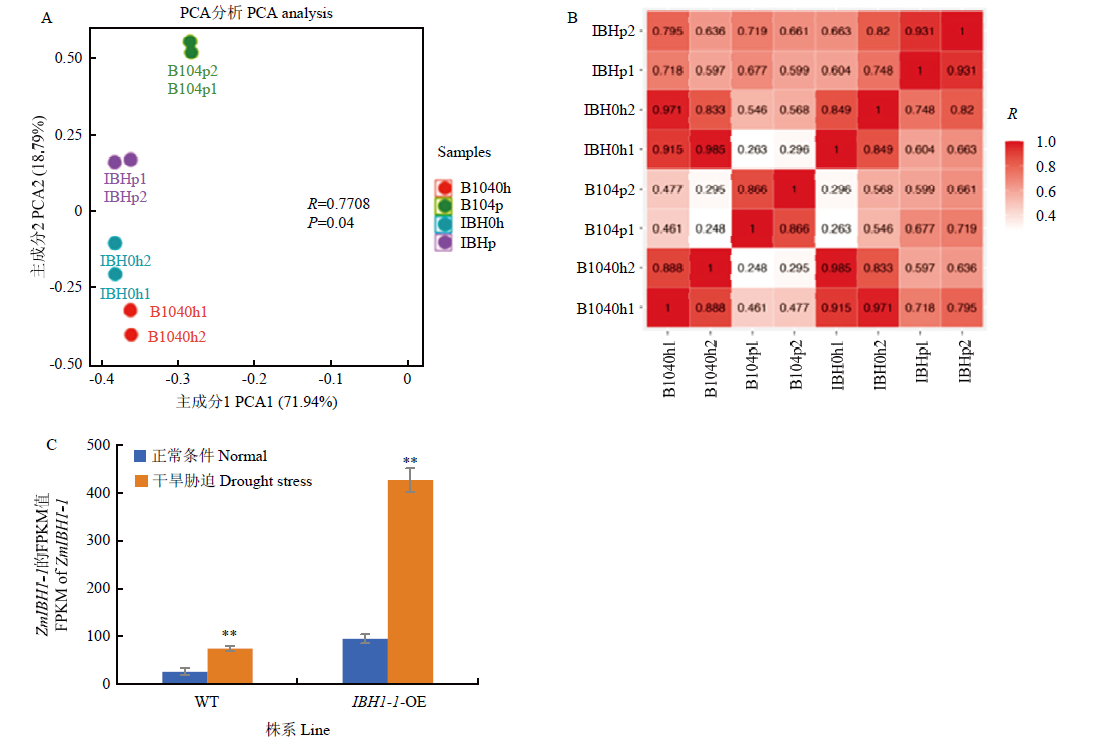
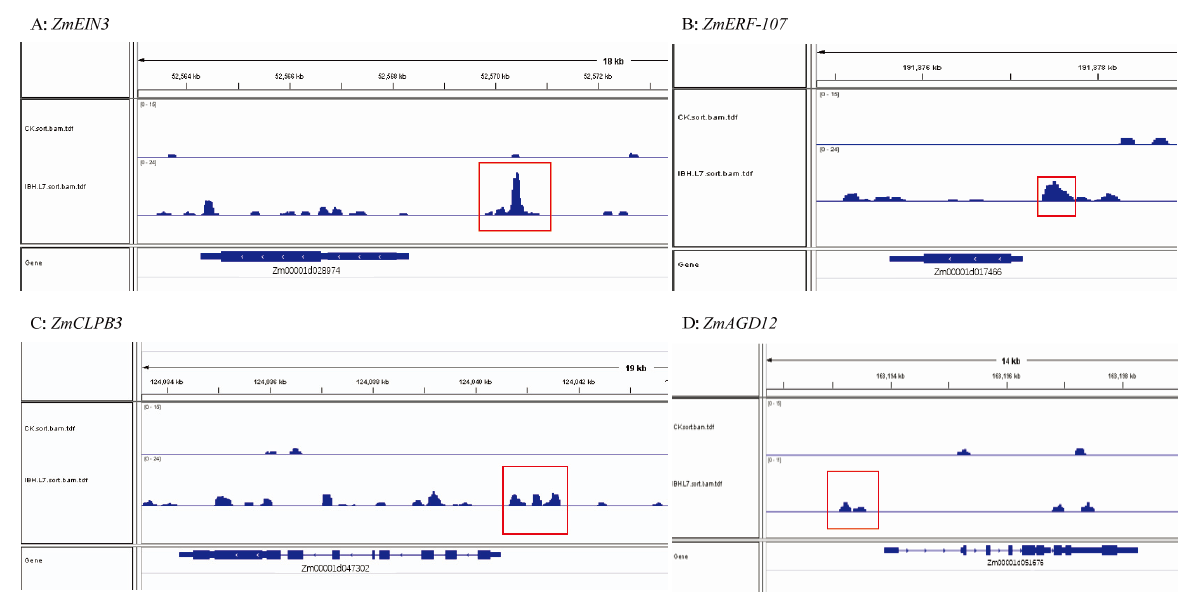
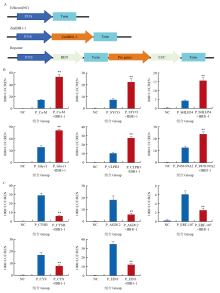
CAO Y, ZENG H, KU L X, REN Z, HAN Y, SU H, DOU D, LIU H, DONG Y, ZHU F. ZmIBH1-1 regulates plant architecture in maize. Journal of Experimental Botany, 2020, 71(10):2943-2955.
doi: 10.1093/jxb/eraa052 |
| [27] |
KENNETH J L, THOMAS D S. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods, 2002, 25:402-408.
doi: 10.1006/meth.2001.1262 |
| [28] | 韩赞平. 玉米种子活力相关性状QTL定位及相关基因的克隆[D]. 郑州: 河南农业大学, 2014. |
| HAN Z P. QTL mapping of seed vigor related traits and related gene cloning in maize[D]. Zhengzhou: Henan Agricultural University, 2014. (in Chinese) | |
| [29] | 姚晓云, 蓝海军, 邓伟, 陈红萍, 罗晨曦, 况震, 罗宗铭, 王记林, 陈大洲. 水稻淡白叶突变体的叶绿素含量测定及农艺性状比较分析. 江西农业学报, 2020, 32(12):12-15. |
| YAO X Y, LAN H J, DENG W, CHEN H P, LUO C X, KUANG Z, LUO Z M, WANG L J, CHEN D Z. Determination of chlorophyll content and comparative analysis of agronomic traits of pale-white- leaf mutant in rice. Acta Agriculturae Jiangxi, 2020, 32(12):12-15. (in Chinese) | |
| [30] | 焦洁. 考马斯亮蓝G-250染色法测定苜蓿中可溶性蛋白含量. 农业工程技术, 2016, 36(17):33-34. |
| JIAO J. Determination of soluble protein content in Alfalfa by Coomassie brilliant blue G-250 staining. Agricultural Engineering Technology, 2016, 36(17):33-34. (in Chinese) | |
| [31] |
YE J. WEGO: A web tool for plotting GO annotations. Nucleic Acids Research, 2006, 34(Web Server issue):W293-W297.
doi: 10.1093/nar/gkl031 |
| [32] |
PIERIK R, SASIDHARAN R, VOESENEK L. Growth control by ethylene: Adjusting phenotypes to the environment. Journal of Plant Growth Regulation, 2007, 26(2):188-200.
doi: 10.1007/s00344-006-0124-4 |
| [33] | LANAHAN M B. The never ripe mutation blocks ethylene perception in tomato. The Plant Cell, 1994, 6(4):521-530. |
| [34] |
GRBI V, Bleecker A B. Ethylene regulates the timing of leaf senescence in Arabidopsis. The Plant Journal, 1995, 8(4):595-602.
doi: 10.1046/j.1365-313X.1995.8040595.x |
| [35] |
LUO J, MA N, PEI H, CHEN J, LI J, GAO J. A DELLA gene, RhGAI1, is a direct target of EIN3 and mediates ethylene-regulated rose petal cell expansion via repressing the expression of RhCesA2. Journal of Experimental Botany, 2013, 64(16):5075-5084.
doi: 10.1093/jxb/ert296 |
| [36] |
HUA J, MEYEROWITZ E M. Ethylene responses are negatively regulated by a receptor gene family in Arabidopsis thaliana. Cell (Cambridge), 1998, 94(2):261-271.
doi: 10.1016/S0092-8674(00)81425-7 |
| [37] |
GUO H W. Paradigms and paradox in the ethylene signaling pathway and interaction network. Molecular Plant, 2011, 4(4):626-634.
doi: 10.1093/mp/ssr042 |
| [38] | OHME-TAKAGI M, SHINSHI H. Ethylene-inducible DNA binding proteins that interact with an ethylene-responsive element. The Plant Cell, 1995, 7(2):173-182. |
| [39] |
HATTORI Y, NAGAI K, FURUKAWA S, SONG XJ, KAWANO R, SAKAKIBARA H, WU J, MATSUMOTO T, YOSHIMURA A, KITANO H, MATSUOKA M, MORI H, ASHIKARI M. The ethylene response factors SNORKEL1 and SNORKEL2 allow rice to adapt to deep water. Nature, 2009, 460(7258):1026-1030.
doi: 10.1038/nature08258 |
| [40] | KNIGHT H. Calcium signaling during abiotic stress in plants. International Review of Cytology-a Survey of Cell Biology, 1999, 195:269-324. |
| [41] |
REDDY A, ALI G S, CELESNIK H, DAY I S. Coping with stresses: Roles of calcium- and calcium/calmodulin-regulated gene expression. The Plant Cell, 2011, 23(6):2010-2032.
doi: 10.1105/tpc.111.084988 |
| [42] |
DUBROVINA A S, KISELEV K V, KHRISTENKO V S, ALEYNOVA O A. VaCPK20, a calcium-dependent protein kinase gene of wild grapevine Vitis amurensis Rupr., mediates cold and drought stress tolerance. Journal of Plant Physiology, 2015, 185:1-12.
doi: 10.1016/j.jplph.2015.05.020 |
| [43] |
ZOU J J, LI X D, RATNASEKERA D, WANG C, LIU W X, SONG L F, ZHANG W Z, WU W H. Arabidopsis CALCIUM-DEPENDENT PROTEIN KINASE8 and CATALASE3 function in abscisic acid-mediated signaling and H2O2 homeostasis in stomatal guard cells under drought stress. The Plant Cell, 2015, 27(5):1445-1460.
doi: 10.1105/tpc.15.00144 |
| [44] |
REN Z, ZHANG D, CAO L, ZHANG W, KU L. Functions and regulatory framework of ZmNST3 in maize under lodging and drought stress. Plant Cell and Environment, 2020, 43(9):2272-2286.
doi: 10.1111/pce.v43.9 |
| [45] |
OH JE, KWON Y, KIM JH, NOH H, HONG SW, LEE H. A dual role for MYB60 in stomatal regulation and root growth of Arabidopsis thaliana under drought stress. Plant Molecular Biology, 2011, 77(1/2):91-103.
doi: 10.1007/s11103-011-9796-7 |
| [46] |
NAKABAYASHI R, YONEKURA-SAKAKIBARA K, URANO K, SUZUKI M, YAMADA Y, NISHIZAWA T, MATSUDA F, KOJIMA M, SAKAKIBARA H, SHINOZAKI K. Enhancement of oxidative and drought tolerance in Arabidopsis by overaccumulation of antioxidant flavonoids. The Plant Journal, 2014, 77(3):367-379.
doi: 10.1111/tpj.2014.77.issue-3 |
| [47] |
DAI X Y, XU Y Y, MA Q B, XU W Y, WANG T, XUE Y B, CHONG K. Overexpression of an R1R2R3 MYB gene, OsMYB3R-2, increases tolerance to freezing, drought, and salt stress in transgenic Arabidopsis. Plant Physiology, 2007, 143(4):1739-1751.
doi: 10.1104/pp.106.094532 |
| [48] |
HONG S, CHEN S, JIANG J, CHEN F, CHEN Y, GU C, LI P, SONG A, ZHU X, GAO H. Heterologous expression of the chrysanthemum R2R3-MYB Transcription Factor CmMYB2 enhances drought and salinity tolerance, increases hypersensitivity to ABA and delays flowering in Arabidopsis thaliana. Molecular Biotechnology, 2012, 51(2):160-173.
doi: 10.1007/s12033-011-9451-1 |
| [49] |
QIN Y, WANG M, TIAN Y, HE W, LU H, XIA G. Over-expression of TaMYB33 encoding a novel wheat MYB transcription factor increases salt and drought tolerance in Arabidopsis. Molecular Biology Reports, 2012, 39(6):7183-7192.
doi: 10.1007/s11033-012-1550-y |
| [50] | FINATTO T, VIANA V E, WOYANN L G, BUSANELLO C, OLIVEIRA A. Can WRKY transcription factors help plants to overcome environmental challenges? Genetics & Molecular Biology, 2018, 41(3):533-544. |
| [1] | CHAI HaiYan,JIA Jiao,BAI Xue,MENG LingMin,ZHANG Wei,JIN Rong,WU HongBin,SU QianFu. Identification of Pathogenic Fusarium spp. Causing Maize Ear Rot and Susceptibility of Some Strains to Fungicides in Jilin Province [J]. Scientia Agricultura Sinica, 2023, 56(1): 64-78. |
| [2] | GU LiDan,LIU Yang,LI FangXiang,CHENG WeiNing. Cloning of Small Heat Shock Protein Gene Hsp21.9 in Sitodiplosis mosellana and Its Expression Characteristics During Diapause and Under Temperature Stresses [J]. Scientia Agricultura Sinica, 2023, 56(1): 79-89. |
| [3] | ZHAO ZhengXin,WANG XiaoYun,TIAN YaJie,WANG Rui,PENG Qing,CAI HuanJie. Effects of Straw Returning and Nitrogen Fertilizer Types on Summer Maize Yield and Soil Ammonia Volatilization Under Future Climate Change [J]. Scientia Agricultura Sinica, 2023, 56(1): 104-117. |
| [4] | ZHANG KeKun,CHEN KeQin,LI WanPing,QIAO HaoRong,ZHANG JunXia,LIU FengZhi,FANG YuLin,WANG HaiBo. Effects of Irrigation Amount on Berry Development and Aroma Components Accumulation of Shine Muscat Grape in Root-Restricted Cultivation [J]. Scientia Agricultura Sinica, 2023, 56(1): 129-143. |
| [5] | HU Sheng,LI YangYang,TANG ZhangLin,LI JiaNa,QU CunMin,LIU LieZhao. Genome-Wide Association Analysis of the Changes in Oil Content and Protein Content Under Drought Stress in Brassica napus L. [J]. Scientia Agricultura Sinica, 2023, 56(1): 17-30. |
| [6] | LI ZhouShuai,DONG Yuan,LI Ting,FENG ZhiQian,DUAN YingXin,YANG MingXian,XU ShuTu,ZHANG XingHua,XUE JiQuan. Genome-Wide Association Analysis of Yield and Combining Ability Based on Maize Hybrid Population [J]. Scientia Agricultura Sinica, 2022, 55(9): 1695-1709. |
| [7] | XIONG WeiYi,XU KaiWei,LIU MingPeng,XIAO Hua,PEI LiZhen,PENG DanDan,CHEN YuanXue. Effects of Different Nitrogen Application Levels on Photosynthetic Characteristics, Nitrogen Use Efficiency and Yield of Spring Maize in Sichuan Province [J]. Scientia Agricultura Sinica, 2022, 55(9): 1735-1748. |
| [8] | LI YiLing,PENG XiHong,CHEN Ping,DU Qing,REN JunBo,YANG XueLi,LEI Lu,YONG TaiWen,YANG WenYu. Effects of Reducing Nitrogen Application on Leaf Stay-Green, Photosynthetic Characteristics and System Yield in Maize-Soybean Relay Strip Intercropping [J]. Scientia Agricultura Sinica, 2022, 55(9): 1749-1762. |
| [9] | QIU YiLei,WU Fan,ZHANG Li,LI HongLiang. Effects of Sublethal Doses of Imidacloprid on the Expression of Neurometabolic Genes in Apis cerana cerana [J]. Scientia Agricultura Sinica, 2022, 55(8): 1685-1694. |
| [10] | MA XiaoYan,YANG Yu,HUANG DongLin,WANG ZhaoHui,GAO YaJun,LI YongGang,LÜ Hui. Annual Nutrients Balance and Economic Return Analysis of Wheat with Fertilizers Reduction and Different Rotations [J]. Scientia Agricultura Sinica, 2022, 55(8): 1589-1603. |
| [11] | LI Qian,QIN YuBo,YIN CaiXia,KONG LiLi,WANG Meng,HOU YunPeng,SUN Bo,ZHAO YinKai,XU Chen,LIU ZhiQuan. Effect of Drip Fertigation Mode on Maize Yield, Nutrient Uptake and Economic Benefit [J]. Scientia Agricultura Sinica, 2022, 55(8): 1604-1616. |
| [12] | ZHANG JiaHua,YANG HengShan,ZHANG YuQin,LI CongFeng,ZHANG RuiFu,TAI JiCheng,ZHOU YangChen. Effects of Different Drip Irrigation Modes on Starch Accumulation and Activities of Starch Synthesis-Related Enzyme of Spring Maize Grain in Northeast China [J]. Scientia Agricultura Sinica, 2022, 55(7): 1332-1345. |
| [13] | TAN XianMing,ZHANG JiaWei,WANG ZhongLin,CHEN JunXu,YANG Feng,YANG WenYu. Prediction of Maize Yield in Relay Strip Intercropping Under Different Water and Nitrogen Conditions Based on PLS [J]. Scientia Agricultura Sinica, 2022, 55(6): 1127-1138. |
| [14] | DONG SangJie,JIANG XiaoChun,WANG LingYu,LIN Rui,QI ZhenYu,YU JingQuan,ZHOU YanHong. Effects of Supplemental Far-Red Light on Growth and Abiotic Stress Tolerance of Pepper Seedlings [J]. Scientia Agricultura Sinica, 2022, 55(6): 1189-1198. |
| [15] | LIU Miao,LIU PengZhao,SHI ZuJiao,WANG XiaoLi,WANG Rui,LI Jun. Critical Nitrogen Dilution Curve and Nitrogen Nutrition Diagnosis of Summer Maize Under Different Nitrogen and Phosphorus Application Rates [J]. Scientia Agricultura Sinica, 2022, 55(5): 932-947. |
|
||