Scientia Agricultura Sinica ›› 2020, Vol. 53 ›› Issue (23): 4777-4790.doi: 10.3864/j.issn.0578-1752.2020.23.005
• PLANT PROTECTION • Previous Articles Next Articles
WANG BaoBao1,GUO Cheng2,SUN SuLi1,XIA YuSheng1,ZHU ZhenDong1,DUAN CanXing1( )
)
| [1] |
王晓鸣, 段灿星 . 玉米病害和病原名称整理及其汉译名称规范化探讨. 中国农业科学, 2020,53(2):288-316.
doi: 10.3864/j.issn.0578-1752.2020.02.006 |
|
WANG X M, DUAN C X . Reorganization of maize disease and causal agent names and disscution on their standardized translation of Chinese names. Scientia Agricultura Sinica, 2020,53(2):288-316. (in Chinese)
doi: 10.3864/j.issn.0578-1752.2020.02.006 |
|
| [2] | 王宝宝, 毕四刚, 肖明纲, 张冬英, 闫强, 张彦彦, 杨树龙, 朱振东, 段灿星 . 黑龙江省玉米穗腐病致病镰孢菌分离鉴定及产毒基因型分析. 草业学报, 2020,29(1):163-174. |
| WANG B B, BI S G, XIAO M G, ZHANG D Y, YAN Q, ZHANG Y Y, YANG S L, ZHU Z D, DUAN C X . Isolation and identification of pathogenic Fusarium spp. causing maize ear rot and analysis of their toxin-producing genotype in Heilongjiang Province. Acta Prataculturae Sinica, 2020,29(1):163-174. (in Chinese) | |
| [3] |
DUAN C X, QIN Z H, YANG Z H, LI W X, SUN S L, ZHU Z D, WANG X M . Identification of pathogenic Fusarium spp. causing maize ear rot and potential mycotoxin production in China. Toxins, 2016,8(6):186.
doi: 10.3390/toxins8060186 |
| [4] |
段灿星, 王晓鸣, 宋凤景, 孙素丽, 周丹妮, 朱振东 . 玉米抗穗腐病研究进展. 中国农业科学, 2015,48(11):2152-2164.
doi: 10.3864/j.issn.0578-1752.2015.11.007 |
|
DUAN C X, WANG X M, SONG F J, SUN S L, ZHOU D N, ZHU Z D . Advances in research on maize resistance to ear rot. Scientia Agricultura Sinica, 2015,48(11):2152-2164. (in Chinese)
doi: 10.3864/j.issn.0578-1752.2015.11.007 |
|
| [5] | 秦子惠, 任旭, 江凯, 武小菲, 杨知还, 王晓鸣 . 我国玉米穗腐病致病镰孢种群及禾谷镰孢复合种的鉴定. 植物保护学报, 2014,41(5):589-596. |
| QIN Z H, REN X, JIANG K, WU X F, YANG Z H, WANG X M . Identification of Fusarium species and F. graminearum species complex causing maize ear rot in China. Journal of Plant Protection, 2014,41(5):589-596. (in Chinese) | |
| [6] | 周丹妮, 王晓鸣, 李丹丹, 杨洋, 陈国康, 段灿星 . 重庆及周边地区玉米穗腐病致病镰孢菌的分离与鉴定. 植物保护学报, 2016,43(5):782-788. |
| ZHOU D N, WANG X M, LI D D, YANG Y, CHEN G K, DUAN C X . Isolation and identification of Fusarium species causing maize ear rot in Chongqing and its vicinity. Journal of Plant Protection, 2016,43(5):782-788. (in Chinese) | |
| [7] |
LI L, QU Q, CAO Z, GUO Z, JIA H, LIU N, WANG Y, DONG J . The relationship analysis on corn stalk rot and ear rot according to Fusarium species and fumonisin contamination in kernels. Toxins, 2019,11(6):320.
doi: 10.3390/toxins11060320 |
| [8] |
ZHU Z, HAO Y, MERGOUM M, BAI G, HUMPHREYS G, CLOUTIER S, XIA X, HE Z . Breeding wheat for resistance to Fusarium head blight in the Global North: China, USA and Canada. The Crop Journal, 2019,7(6):730-738.
doi: 10.1016/j.cj.2019.06.003 |
| [9] | 支叶, 孙菲菲, 孙素丽, 段灿星, 朱振东 . 黑龙江省大豆根腐病菌锐顶镰孢鉴定. 中国油料作物学报, 2014,36(6):789-793. |
| ZHI Y, SUN F F, SUN S L, DUAN C X, ZHU Z D . Identification of Fusarium acuminatum causing soybean root rot in Heilongjiang Province. Chinese Journal of Oil Crop Sciences, 2014,36(6):789-793. (in Chinese) | |
| [10] |
DONG F, ZHANG X, XU J, SHI J, LEE Y W, CHEN X, LI Y, MOKOENA M P, OLANIRAN A O . Analysis of Fusarium graminearum species complex from freshly harvested rice in Jiangsu Province (China). Plant Disease, 2020,104(8): DOI: 10.1094/PDIS- 01-20-0084-RE.
doi: 10.1094/PDIS-11-19-2473-SR pmid: 32525450 |
| [11] |
GROMADZKA K, BLASZCZYK L, CHELKOWSKI J, WASKIEWICZ A . Occurrence of mycotoxigenic Fusarium species and competitive fungi on preharvest maize ear rot in Poland. Toxins, 2019,11(4):224.
doi: 10.3390/toxins11040224 |
| [12] |
MIEDANER T, GWIAZDOWSKA D, WASKIEWICZ A . Editorial: Management of Fusarium species and their mycotoxins in cereal food and feed. Frontiers in Microbiology, 2017,8:1543.
doi: 10.3389/fmicb.2017.01543 pmid: 28861058 |
| [13] |
GAIKPA D S, MIEDANER T . Genomics-assisted breeding for ear rot resistances and reduced mycotoxin contamination in maize: methods, advances and prospects. Theoretical and Applied Genetics, 2019,132(10):2721-2739.
doi: 10.1007/s00122-019-03412-2 pmid: 31440772 |
| [14] |
GAI X T, XUAN Y H, GAO Z G . Diversity and pathogenicity of Fusarium graminearum species complex from corn stalk and ear rot strains in northeast China. Plant Pathology, 2017,66(8):1267-1275.
doi: 10.1111/ppa.2017.66.issue-8 |
| [15] |
马红霞, 孙华, 郭宁, 张海剑, 石洁, 常佳迎 . 禾谷镰孢复合种毒素化学型及遗传多样性分析. 中国农业科学, 2018,51(1):82-95.
doi: 10.3864/j.issn.0578-1752.2018.01.008 |
|
MA H X, SUN H, GUO N, ZHANG H J, SHI J, CHANG J Y . Analysis of toxigenic chemotype and genetic diversity of the Fusarium graminearum species complex. Scientia Agricultura Sinica, 2018,51(1):82-95. (in Chinese)
doi: 10.3864/j.issn.0578-1752.2018.01.008 |
|
| [16] |
YERKOVICH N, FUMERO M V, CANTORO R, PALAZZINI J M, CHULZE S N . Population structure and genetic diversity of Fusarium graminearum sensu stricto, the main wheat pathogen producing Fusarium head blight in Argentina. European Journal of Plant Pathology, 2020,156(2):635-646.
doi: 10.1007/s10658-019-01913-w |
| [17] |
SNEIDERIS D, IVANAUSKAS A, SUPRONIENE S, KADZIENE G, SAKALAUSKAS S . Genetic diversity of Fusarium graminearum isolated from weeds. European Journal of Plant Pathology, 2019,153(2):639-643.
doi: 10.1007/s10658-018-1543-3 |
| [18] | 任旭 . 我国玉米穗腐病主要致病镰孢菌多样性研究[D]. 北京: 中国农业科学院, 2011. |
| REN X . Diversity analyses of Fusarium spp., the main causal agents of maize ear rot in China[D]. Beijing: Chinese Academy of Agricultural Sciences, 2011. ( in Chinese) | |
| [19] | 李新凤, 王建明, 姜晓东, 郝晓娟, 张祖维, 田宏先 . 拟轮枝镰孢菌EST-SSR信息分析与标记开发. 植物保护学报, 2018,45(4):819-826. |
| LI X F, WANG J M, JIANG X D, HAO X J, ZHANG Z W, TIAN H X . EST-SSR information analysis and marker development for genetic diversity analysis of Fusarium verticillioides. Journal of Plant Protection, 2018,45(4):819-826. (in Chinese) | |
| [20] |
SCOTT J B, CHAKRABORTY S . Identification of 11 polymorphic simple sequence repeat loci in the phytopathogenic fungus Fusarium pseudograminearum as a tool for genetic studies. Molecular Ecology Resources, 2008,8(3):628-630.
doi: 10.1111/j.1471-8286.2007.02025.x pmid: 21585853 |
| [21] |
SUHA H, GALE L R, KISTLER H C . Development of VNTR markers for two Fusarium graminearum clade species. Molecular Ecology Notes, 2004,4(3):468-470.
doi: 10.1111/men.2004.4.issue-3 |
| [22] |
O’DONNELL K, WARD T J, ABERRA D, KISTLER H C, AOKI T, ORWIG N, KIMURA M, BJORNSTAD A, KLEMSDAL S S . Multilocus genotyping and molecular phylogenetics resolve a novel head blight pathogen within the Fusarium graminearum species complex from Ethiopia. Fungal Genetics and Biology, 2008,45(11):1514-1522.
doi: 10.1016/j.fgb.2008.09.002 |
| [23] |
HUBKA V, KOLARIK M . β-tubulin paralogue tubC is frequently misidentified as the benA gene in Aspergillus section Nigri taxonomy: Primer specificity testing and taxonomic consequences. Persoonia - Molecular Phylogeny and Evolution of Fungi, 2012,29:1-10.
doi: 10.3767/003158512X658123 |
| [24] |
LOFGREN L A, LEBLANC N R, CERTANO A K, NACHTIGALL J, LABINE K M, RIDDLE J, BROZ K, DONG Y, BETHAN B, KAFER C W, KISTLER H C . Fusarium graminearum: Pathogen or endophyte of North American grasses?. New Phytologist, 2018,217(3):1203-1212.
doi: 10.1111/nph.14894 pmid: 29160900 |
| [25] |
WANG J H, LI H P, QU B, ZHANG J B, HUANG T, CHEN F F, LIAO Y C . Development of a generic PCR detection of 3- acetyldeoxynivalenol-, 15-acetyldeoxynivalenol- and nivalenol- chemotypes of Fusarium graminearum clade. International Journal of Molecular Sciences, 2008,9(12):2495-2504.
doi: 10.3390/ijms9122495 pmid: 19330088 |
| [26] |
WAALWIJK C, KASTELEIN P, DE VRIES I, KERÉNYI Z, VAN DER LEE T, HESSELINK T, KÖHL J, KEMA G . Major changes in Fusarium spp. in wheat in the Netherlands. European Journal of Plant Pathology, 2003,109(7):743-754.
doi: 10.1023/A:1026086510156 |
| [27] |
JENNINGS P, COATES M E, TURNER J A, CHANDLER E A, NICHOLSON P . Determination of deoxynivalenol and nivalenol chemotypes of Fusarium culmorum isolates from England and Wales by PCR assay. Plant Pathology, 2004,53(2):182-190.
doi: 10.1111/ppa.2004.53.issue-2 |
| [28] | DUAN C X, WANG B B, SUN F F, YANG Z H, ZHU Z D, WANG X M . Occurrence of maize ear rot caused by Fusarium fujikuroi in China. Plant Disease, 2020,104(2):587. |
| [29] |
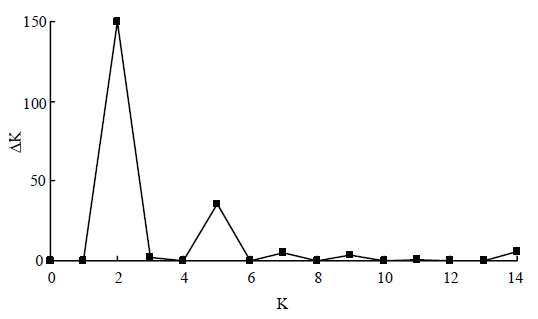
EVANNO G, REGNAUT S, GOUDET J . Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Molecular Ecology, 2005,14(8):2611-2620.
doi: 10.1111/j.1365-294X.2005.02553.x pmid: 15969739 |
| [30] |
BEUKES I, ROSE L J, VAN COLLER G J, VILJOEN A . Disease development and mycotoxin production by the Fusarium graminearum species complex associated with South African maize and wheat. European Journal of Plant Pathology, 2018,150:893-910.
doi: 10.1007/s10658-017-1331-5 |
| [31] |
COAN M, SENHORINHO H J, PINTO R J, SCAPIM C A, TESSMANN D J, WILLIAMS W P, WARBURTON M L . Genome-wide association study of resistance to ear rot by Fusarium verticillioides in a tropical field maize and popcorn core collection. Crop Science, 2018,58(2):564-578.
doi: 10.2135/cropsci2017.05.0322 |
| [32] |
AAMOT H U, WARD T J, BRODAL G, VRALSTAD T, LARSEN G B, KLEMSDAL S S, ELAMEEN A, UHLIG S, HOFGAARD I S . Genetic and phenotypic diversity within the Fusarium graminearum species complex in Norway. European Journal of Plant Pathology, 2015,142(3):501-519.
doi: 10.1007/s10658-015-0629-4 |
| [33] |
WARD T J, CLEAR R M, ROONEY A P, O’DONNELL K, GABA D, PATRICK S, STARKEY D E, GILBERT J, GEISER D M, NOWICKI T W . An adaptive evolutionary shift in Fusarium head blight pathogen populations is driving the rapid spread of more toxigenic Fusarium graminearum in North America. Fungal Genetics and Biology, 2008,45(4):473-484.
doi: 10.1016/j.fgb.2007.10.003 |
| [34] | 李伟, 胡迎春, 陈莹, 张爱香, 陈怀谷 . 长江流域禾谷镰孢菌群部分菌株系统发育学、产毒素化学型及致病力研究. 菌物学报, 2010,29(1):51-58. |
| LI W, HU Y C, CHEN Y, ZHANG A X, CHEN H G . Phylogenetic analysis, chemotype diversity, and pathogenicity of the Fusarium graminearum clade in the Yangtze basin. Mycosystema, 2010,29(1):51-58. (in Chinese) | |
| [35] |
杜青, 唐照磊, 李石初, 上官玲玲, 李华娇, 段灿星 . 广西玉米穗腐病致病镰孢种群构成与毒素化学型分析. 中国农业科学, 2019,52(11):1895-1907.
doi: 10.3864/j.issn.0578-1752.2019.11.005 |
|
DU Q, TANG Z L, LI S C, SHANGGUAN L L, DUAN C X . Composition of Fusarium species causing maize ear rot and analysis of toxigenic chemotype in Guangxi. Scientia Agricultura Sinica, 2019,52(11):1895-1907. (in Chinese)
doi: 10.3864/j.issn.0578-1752.2019.11.005 |
|
| [36] |
LAURENT B, MOINARD M, SPATARO C, PONTS N, BARREAU C, FOULONGNE-ORIOL M . Landscape of genomic diversity and host adaptation in Fusarium graminearum. BMC Genomics, 2017,18:203.
doi: 10.1186/s12864-017-3524-x pmid: 28231761 |
| [1] | DONG Yu, WU Qian, FENG Xuan, ZHENG YinYing, CUI BaiMing. A Novel Plasmid pEA60 of Erwinia amylovora Enhances the Pathogenicity of Strains by Regulating the Synthesis of Virulence Factors [J]. Scientia Agricultura Sinica, 2026, 59(5): 996-1007. |
| [2] | HE ZhiLin, SUN CuiXia, YUE HongLi, TAN YueXia, ZHANG YaoHai, WANG FuSheng, LIU SiTao, JIANG Dong. Genetic Diversity Analysis and GWAS of Alloocimene Based on Resequencing of Citron, Lemon Germplasm Resources [J]. Scientia Agricultura Sinica, 2026, 59(2): 386-401. |
| [3] | CONG QiQi, ZHANG JingYi, MENG XiangLong, DAI PengBo, LI Bo, HU TongLe, WANG ShuTong, CAO KeQiang, WANG YaNan. Identification of Hypovirus in Apple Ring Rot Fungus Botryosphaeria dothidea and Detection of Virus-Carrying Status in China [J]. Scientia Agricultura Sinica, 2025, 58(3): 478-492. |
| [4] | TONG ZhaoYang, LIU WenHua, ZHANG GuoXin, DONG ChunYan, ZHANG YanXia, XU XiaoWei, HE Dong, LIU HeChun, LI Yang, WANG FengTao, FENG Jing, YAO XiaoBo, LIU MeiJin, LIN RuiMing. The Relationship Between Occurrence of Hulless Barley Ear Rot and Population Migration of Grass Mite (Siteroptes spp.) [J]. Scientia Agricultura Sinica, 2025, 58(3): 493-506. |
| [5] | XU YuJuan, ZHANG Jie, WANG TianYi, CHEN HaoYang, ZHAO JiaJia, WU BangBang, HAO YuQiong, LI XiaoHua, ZHENG XingWei, ZUO JingJing, ZHENG Jun. Identification of Glu-A3 and Glu-B3 of Low-Molecular-Weight Glutenin in Shanxi Wheat and Its Effect on Quality [J]. Scientia Agricultura Sinica, 2025, 58(24): 5110-5127. |
| [6] | CHEN CaiJin, MA Lin, JIANG QingXue, LIU JinHui, MIAO Tong, ZHANG ZhiPeng, MENG Xiang, MA XiaoRan, ZHOU XinYue, ZHANG Jian, LIU WenHui, WANG XueMin. Genetic Diversity Analysis of Phenotypic Traits of 244 Forage Oat Germplasm Resources [J]. Scientia Agricultura Sinica, 2025, 58(23): 4825-4836. |
| [7] | WEI YiMin, ZHOU MeiLiang, TANG Yu. Origin, Evolution and Spread of Crop Buckwheat [J]. Scientia Agricultura Sinica, 2025, 58(21): 4305-4316. |
| [8] | LIU XiaoXu, ZHONG ZeXin, QIU JiaRen, YANG ChunXiao, ZHANG YongJun, XIE Wen, ZHANG YouJun, PAN HuiPeng. GENETIC DIVERSITY OF MTCO1 IN DIFFERENT GEOGRAPHICAL POPULATIONS OF MEGALUROTHRIPS USITATUS [J]. Scientia Agricultura Sinica, 2025, 58(21): 4361-4371. |
| [9] | GUO MengZe, ZHANG Lei, SUN PingPing, JIANG Biao, YAN JinQiang, LI ZhengNan. Molecular Characterization and Evolutionary Dynamics of Tomato Leaf Curl New Delhi Virus Isolate from Wax Gourd (Benincasa hispida) in Guangdong [J]. Scientia Agricultura Sinica, 2025, 58(19): 3890-3904. |
| [10] | LIU PengPeng, LI JiangBo, XU HongJun, NIE YingBin, HAN XinNian, KONG DeZhen, SANG Wei. Genetic Diversity Analysis of Protein Fractions and Quality in Xinjiang Winter Wheat Cultivar Resources [J]. Scientia Agricultura Sinica, 2025, 58(15): 2948-2959. |
| [11] | YANG WenJuan, GAO JiaCheng, WANG YanTing, LI Yan, GUO Ming, WANG JunCheng, MENG YaXiong, WANG HuaJun, SI ErJing. Function of Effector Pg00778 Regulation on the Pathogenicity of Pyrenophora graminea to Barley [J]. Scientia Agricultura Sinica, 2025, 58(15): 3020-3035. |
| [12] | WANG Hui, DING BaoPeng, LI YuXian, REN QuanRu, ZHOU Hai, ZHAO JunLiang, HU HaiFei. Research Progress and Prospects on Crop Pan-Genomics [J]. Scientia Agricultura Sinica, 2025, 58(11): 2045-2061. |
| [13] | DONG ZaiFang, DING TengTeng, SHAN YiXuan, LI HongLian, CHEN LinLin, XING XiaoPing. Autophagy-Related Gene FpAtg3 Involves in Growth and Pathogenicity of Fusarium pseudograminearum [J]. Scientia Agricultura Sinica, 2024, 57(6): 1080-1090. |
| [14] | ZHANG AiHong, YANG Fei, ZHAO YuanYe, ZHAO YiHan, DI DianPing, MIAO HongQin. Pathogenicity and Epidemic Risk of Barley Yellow Striate Mosaic Virus [J]. Scientia Agricultura Sinica, 2024, 57(23): 4686-4697. |
| [15] | LI Pei, HE ZhiLin, TAN YueXia, ZHAO WanTong, FENG JinYing, CHEN GuiHu, YAN Chi, WANG ZiHao, HUANG Ping, JIANG Dong. Genetic Diversity Analysis of Mandarin and Excellent Germplasm Screening Based on Whole-Genome Resequencing Data and Phenotypic Traits [J]. Scientia Agricultura Sinica, 2024, 57(23): 4761-4773. |
|
||