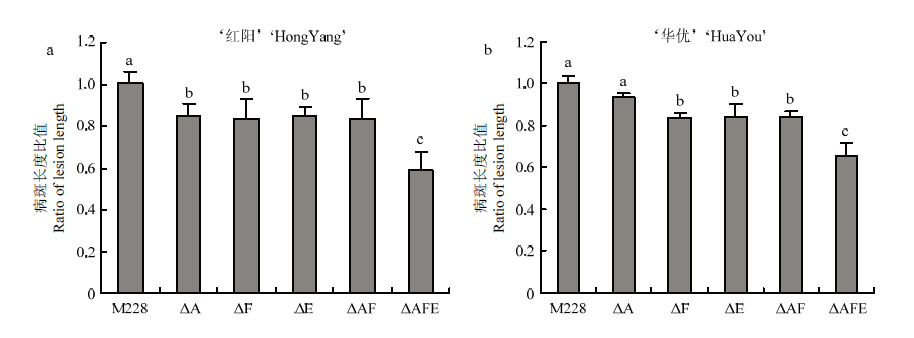
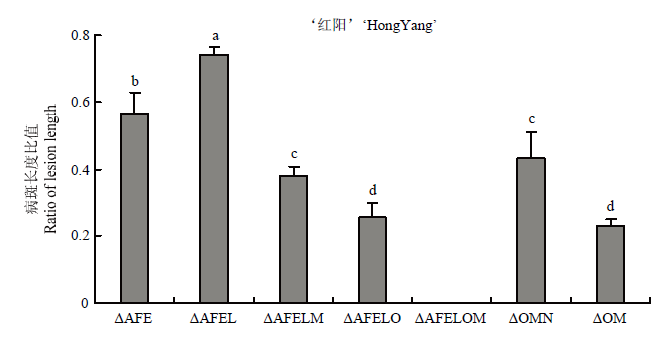
Scientia Agricultura Sinica ›› 2022, Vol. 55 ›› Issue (3): 503-513.doi: 10.3864/j.issn.0578-1752.2022.03.007
• PLANT PROTECTION • Previous Articles Next Articles
ZHANG JinLong( ),ZHAO ZhiBo,LIU Wei,HUANG LiLi()
),ZHAO ZhiBo,LIU Wei,HUANG LiLi()
| [1] | 秦虎强, 高小宁, 赵志博, 朱穗层, 李建民, 黄丽丽. 陕西猕猴桃细菌性溃疡病田间发生动态和规律. 植物保护学报, 2013, 40(3): 225-230. |
| QIN H Q, GAO X N, ZHAO Z B, ZHU H C, LI J M, HUANG L L.The prevalence dynamics and rules of bacterial canker of kiwifruit in Shaanxi. Acta Phytophylacica Sinica, 2013, 40(3): 225-230. (in Chinese) | |
| [2] | 高小宁, 赵志博, 黄其玲, 秦虎强, 黄丽丽. 猕猴桃细菌性溃疡病研究进展. 果树学报, 2012, 29(2): 262-268. |
| GAO X N, ZHAO Z B, HUANG Q L, QIN H Q, HUANG L L.Advances in research on bacterial canker of kiwifruit. Journal of Fruit Science, 2012, 29(2): 262-268. (in Chinese) | |
| [3] |
ZHAO Z, CHEN J, GAO X, ZHANG D, ZHANG J, WEN J, QIN H, GUO M, HUANG L.Comparative genomics reveal pathogenicity- related loci in Pseudomonas syringae pv. actinidiae biovar 3. Molecular Plant Pathology, 2019, 20(7): 923-942.
doi: 10.1111/mpp.2019.20.issue-7 |
| [4] | 孙思, 牛建军, 王岱. 细菌三型分泌系统效应蛋白转运的研究进展. 微生物学报, 2017, 57(10): 1452-1460. |
| SUN S, NIU J J, WANG D.Advances in studies of translocation of effector by bacterial type 3 secretion system. Acta Microbiologica Sinica, 2017, 57(10): 1452-1460. (in Chinese) | |
| [5] |
LINDEBERG M, CUNNAC S, COLLMER A.Pseudomonas syringae type III effector repertoires: Last words in endless arguments. Trends in Microbiology, 2012, 20(4): 199-208.
doi: 10.1016/j.tim.2012.01.003 |
| [6] |
DOS SANTOS A M P,FERRARI R G, CONTE-JUNIOR C A. Type three secretion system in Salmonella typhimurium: The key to infection. Genes and Genomics, 2020, 42(5): 495-506.
doi: 10.1007/s13258-020-00918-8 |
| [7] | 朱秀秀, 高必达, 赵廷昌, 张月娟. 植物病原细菌Ⅲ型分泌系统及Pseudomonas syringae pv. tomato的信号分子分泌研究进展. 湖南农业科学, 2009(2): 19-22. |
| ZHU X X, GAO B D, ZHAO Y C, ZHANG Y J.Research progress of type III secretory system of plant pathogenic bacteria and signal molecule secretion of Pseudomonas syringae pv. tomato. Hunan Agricultural Sciences, 2009(2): 19-22. (in Chinese) | |
| [8] |
MARLOVITS T C, KUBORI T, LARA-TEJERO M, THOMAS D, UNGER V M, GALAN J E.Assembly of the inner rod determines needle length in the type III secretion injectisome. Nature, 2006, 441(7093): 637-640.
doi: 10.1038/nature04822 |
| [9] |
MACHO A P, ZIPFEL C.Plant PRRs and the activation of innate immune signaling. Molecular Cell, 2014, 54(2): 263-272.
doi: 10.1016/j.molcel.2014.03.028 |
| [10] | 温晶. 猕猴桃溃疡病菌Ⅲ型效应蛋白的筛选及效应蛋白HopX3功能的初步研究[D]. 杨凌: 西北农林科技大学, 2016. |
| WEN J.Identification of Psa type III effectors and preliminary analysis of effector HopX3 in pathogenicity[D]. Yangling: Northwest A&F University, 2016. (in Chinese) | |
| [11] | BALTRUS D A, NISHIMURA M T, ROMANCHUK A, CHANG J H,MUKHTAR M S, CHERKIS K, ROACH J, GRANT S R, JONES C D, DANGL J L.Dynamic evolution of pathogenicity revealed by sequencing and comparative genomics of 19 Pseudomonas syringae isolates. PLoS Pathogens, 2011, 7(7): e1002132. |
| [12] | CUNNAC S, CHAKRAVARTHY S, KVITKO B H, RUSSELL A B, MARTIN G B, COLLMER A.Genetic disassembly and combinatorial reassembly identify a minimal functional repertoire of type III effectors in Pseudomonas syringae. Proceedings of the National Academy of Sciences of the United States of America, 2011, 108(7): 2975-2980. |
| [13] |
KVITKO B H, PARK D H, VELASQUEZ A C, WEI C F, RUSSELL A B, MARTIN G B, SCHNEIDER D J, COLLMER A.Deletions in the repertoire of Pseudomonas syringae pv. tomato DC3000 type III secretion effector genes reveal functional overlap among effectors. PLoS Pathogens, 2009, 5(4): e1000388.
doi: 10.1371/journal.ppat.1000388 |
| [14] |
TAMPAKAKI A P, SKANDILIS N, GAZI A D, BASTAKI M N, PANAGIOTIS F S, CHAROVA S N, KOKKINIDIS M, PANOPOULOS N J.Playing the “Harp”: Evolution of our understanding of hrp/hrc genes 1. Annual Review of Phytopathology, 2010, 48: 347-370.
doi: 10.1146/phyto.2010.48.issue-1 |
| [15] |
CHOI S, JAYARAMAN J, SEGONZAC C, PARK H J, PARK H, HAN S W, SOHN K H.Pseudomonas syringae pv. actinidiae type III effectors localized at multiple cellular compartments activate or suppress innate immune responses in Nicotiana benthamiana. Frontiers in Plant Science, 2017, 8: 2157.
doi: 10.3389/fpls.2017.02157 |
| [16] |
JAYARAMAN J, CHOI S, PROKCHORCHIK M, CHOI D S, SPIANDORE A, RIKKERINK E H, TEMPLETON M D, SEGONZAC C, SOHN K H.A bacterial acetyltransferase triggers immunity in Arabidopsis thaliana independent of hypersensitive response. Scientific Reports, 2017, 7(1): 3557.
doi: 10.1038/s41598-017-03704-x |
| [17] |
CHOI S, JAYARAMAN J, SOHN K H.Arabidopsis thaliana SOBER1 (SUPPRESSOR OF AVRBST-ELICITED RESISTANCE 1) suppresses plant immunity triggered by multiple bacterial acetyltransferase effectors. New Phytologist, 2018, 219(1): 324-335.
doi: 10.1111/nph.2018.219.issue-1 |
| [18] | YOON M, RIKKERINK E H A. Rpa1 mediates an immune response to avrRpm1Psa and confers resistance against Pseudomonas syringae pv. actinidiae. The Plant Journal, 2020, 102(4): 688-702. |
| [19] |
JAYARAMAN J, YOON M, APPLEGATE E R, STROUD E A, TEMPLETON M D.AvrE1 and HopR1 from Pseudomonas syringae pv. actinidiae are additively required for full virulence on kiwifruit. Molecular Plant Pathology, 2020, 21(11): 1467-1480.
doi: 10.1111/mpp.12989 |
| [20] | 赵志博. 猕猴桃细菌性溃疡病菌群体结构与致病机制研究[D]. 杨凌: 西北农林科技大学, 2016. |
| ZHAO Z B.Population composition and pathogenetic mechanism in Psuesdomonas syringae pv. actinidiae[D]. Yangling: Northwest A&F University, 2016. (in Chinese) | |
| [21] |
WANG K, KANG L, ANAND A, LAZAROVITS G, MYSORE K S.Monitoring in planta bacterial infection at both cellular and whole-plant levels using the green fluorescent protein variant GFPuv. New Phytologist, 2007, 174(1): 212-223.
doi: 10.1111/nph.2007.174.issue-1 |
| [22] | KVITKO B H, COLLMER A.Construction of Pseudomonas syringae pv. tomato DC3000 mutant and polymutant strains//Plant Immunity. Methods and Protocols, 2011, 712: 109-128. |
| [23] |
SAWADA H, FUJIKAWA T.Genetic diversity of Pseudomonas syringae pv. actinidiae, pathogen of kiwifruit bacterial canker. Plant Pathology, 2019, 68(7): 1235-1248.
doi: 10.1111/ppa.v68.7 |
| [24] |
XIN X F, NOMURA K, AUNG K, VELASQUEZ A C, YAO J, BOUTROT F, CHANG J H, ZIPFEL C, HE S Y.Bacteria establish an aqueous living space in plants crucial for virulence. Nature, 2016, 539(7630): 524-529.
doi: 10.1038/nature20166 |
| [25] |
JIN L, HAM J H, HAGE R, ZHAO W, SOTO-HERNANDEZ J, LEE S Y, PAEK S M, KIM M G, BOONE C, COPLIN D L, MACKEY D.Direct and indirect targeting of PP2A by conserved bacterial type-III effector proteins. PLoS Pathogens, 2016, 12(5): e1005609.
doi: 10.1371/journal.ppat.1005609 |
| [26] |
DEGRAVE A, SIAMER S, BOUREAU T, BARNY M A.The AvrE superfamily: Ancestral type III effectors involved in suppression of pathogen-associated molecular pattern-triggered immunity. Molecular Plant Pathology, 2015, 16(8): 899-905.
doi: 10.1111/mpp.2015.16.issue-8 |
| [27] | PALACE S G, PROULX M K, SZABADY R L, GOGUEN J D.Gain-of-function analysis reveals important virulence roles for the Yersinia pestis type III secretion system effectors YopJ, YopT, and YpkA. Infection and Immunity, 2018, 86(9): e00318-18. |
| [28] |
ÜSTÜN S, KÖNIG P, GUTTMAN D S, BÖRNKE F. HopZ4 from Pseudomonas syringae, a member of the HopZ type III effector family from the YopJ superfamily, inhibits the proteasome in plants. Molecular Plant-Microbe Interactions, 2014, 27(7): 611-623.
doi: 10.1094/MPMI-12-13-0363-R |
| [29] | LEWIS J D, LEE A H Y,HASSAN J A,WAN J,HURLEY B,JHINGREE J R,WANG P W,LO T,YOUN J Y,GUTTMAN D S,DESVEAUX D. The Arabidopsis ZED1 pseudokinase is required for ZAR1-mediated immunity induced by the Pseudomonas syringae type III effector HopZ1a. Proceedings of the National Academy of Sciences of the United States of America, 2013, 110(46): 18722-18727. |
| [30] |
LEWIS J D, LEE A, MA W B, ZHOU H B, GUTTMAN D S, DESVEAUX D.The YopJ superfamily in plant-associated bacteria. Molecular Plant Pathology, 2011, 12(9): 928-937.
doi: 10.1111/j.1364-3703.2011.00719.x |
| [31] |
ZHOU H B, LIN J A, JOHNSON A, MORGAN R L, ZHONG W W, MA W B.Pseudomonas syringae type III effector HopZ1 targets a host enzyme to suppress isoflavone biosynthesis and promote infection in soybean. Cell Host and Microbe, 2011, 9(3): 177-186.
doi: 10.1016/j.chom.2011.02.007 |
| [32] |
MACHO A P, GUIDOT A, BARBERIS P, BEUZON C R, GENIN S.A competitive index assay identifies several Ralstonia solanacearum type III effector mutant strains with reduced fitness in host plants. Molecular Plant-Microbe Interactions, 2010, 23(9): 1197-1205.
doi: 10.1094/MPMI-23-9-1197 |
| [33] |
BARTETZKO V, SONNEWALD S, VOGEL F, HARTNER K, STADLER R, HAMMES U Z, BORNKE F.The Xanthomonas campestris pv. vesicatoria type III effector protein XopJ inhibits protein secretion: Evidence for interference with cell wall-associated defense eesponses. Molecular Plant-Microbe Interactions, 2009, 22(6): 655-664.
doi: 10.1094/MPMI-22-6-0655 |
| [34] |
CHEN H, HU Y, QIN K Y, YANG X Z, JIA Z J, LI Q, CHEN H B, YANG H.A serological approach for the identification of the effector hopz5 of Pseudomonas syringae pv. actinidiae: A tool for the rapid immunodetection of kiwifruit bacterial canker. Journal of Plant Pathology, 2018, 100(2): 171-177.
doi: 10.1007/s42161-018-0041-y |
| [35] |
KRAUS C M, MUNKVOLD K R, MARTIN G B.Natural variation in tomato reveals differences in the recognition of AvrPto and AvrPtoB effectors from Pseudomonas syringae. Molecular Plant, 2016, 9(5): 639-649.
doi: 10.1016/j.molp.2016.03.001 |
| [36] |
KIM M G, GENG X, LEE S Y, MACKEY D.The Pseudomonas syringae type III effector AvrRpm1 induces significant defenses by activating the Arabidopsis nucleotide-binding leucine-rich repeat protein RPS2. The Plant Journal, 2009, 57(4): 645-653.
doi: 10.1111/tpj.2009.57.issue-4 |
| [1] | HUANG JiaQuan,LI Li,WU FengNian,ZHENG Zheng,DENG XiaoLing. Proliferation of Two Types Prophage of ‘Candidatus Liberibacter asiaticus’ in Diaphorina citri and their Pathogenicity [J]. Scientia Agricultura Sinica, 2022, 55(4): 719-728. |
| [2] | YANG ShiMan, XU ChengZhi, XU BangFeng, WU YunPu, JIA YunHui, QIAO ChuanLing, CHEN HuaLan. Amino Acid of 225 in the HA Protein Affects the Pathogenicities of H1N1 Subtype Swine Influenza Viruses [J]. Scientia Agricultura Sinica, 2022, 55(4): 816-824. |
| [3] | LI ZhengGang,TANG YaFei,SHE XiaoMan,YU Lin,LAN GuoBing,HE ZiFu. Molecular Characteristics and Pathogenicity Analysis of Youcai Mosaic Virus Guangdong Isolate Infecting Radish [J]. Scientia Agricultura Sinica, 2022, 55(14): 2752-2761. |
| [4] | ZHANG ChengQi,LIAO LuLu,QI YongXia,DING KeJian,CHEN Li. Functional Analysis of the Nucleoporin Gene FgNup42 in Fusarium graminearium [J]. Scientia Agricultura Sinica, 2021, 54(9): 1894-1903. |
| [5] | HanXi LIU,Hao LÜ,GuangYu GUO,DongXu LIU,Yan SHI,ZhiJun SUN,ZeXin ZHANG,YanJiao ZHANG,YingNan WEN,JieQi WANG,ChunYan LIU,QingShan CHEN,DaWei XIN,JinHui WANG. Effect of rhcN Gene Mutation on Nodulation Ability of Soybean Rhizobium HH103 [J]. Scientia Agricultura Sinica, 2021, 54(6): 1104-1111. |
| [6] | CAO YuHan,LI ZiTeng,ZHANG JingYi,ZHANG JingNa,HU TongLe,WANG ShuTong,WANG YaNan,CAO KeQiang. Analysis of dsRNA Carried by Alternaria alternata f. sp. mali in China and Identification of a dsRNA Virus [J]. Scientia Agricultura Sinica, 2021, 54(22): 4787-4799. |
| [7] | ZHANG Li,TANG YaFei,LI ZhengGang,YU Lin,LAN GuoBing,SHE XiaoMan,HE ZiFu. Molecular Characteristic of Squash Leaf Curl China Virus (SLCCNV) Infecting Cucurbitaceae Crops in Guangdong Province [J]. Scientia Agricultura Sinica, 2021, 54(19): 4097-4109. |
| [8] | ZHAO JingYa,XIA HuiQing,PENG MengYa,FAN Zhuo,YIN Yue,XU SaiBo,ZHANG Nan,CHEN WenBo,CHEN LinLin. Identification and Functional Analysis of Transcription Factors FpAPSES in Fusarium pseudograminearum [J]. Scientia Agricultura Sinica, 2021, 54(16): 3428-3439. |
| [9] | ZHENG XinShi,SHANG PengXiang,LI JingYuan,DING XinLun,WU ZuJian,ZHANG Jie. Effects of Proteins Encoded by “C4 ORFs” of Cotton Leaf Curl Multan Virus on Viral Pathogenicity [J]. Scientia Agricultura Sinica, 2021, 54(10): 2095-2104. |
| [10] | JiaYing CHANG,ShuSen LIU,Jie SHI,Ning GUO,HaiJian ZHANG,HongXia MA,ChunFeng YANG. Pathogenicity and Genetic Diversity of Bipolaria maydis in Sanya, Hainan and Huang-Huai-Hai Region [J]. Scientia Agricultura Sinica, 2020, 53(6): 1154-1165. |
| [11] | LI ZhengGang,NONG Yuan,TANG YaFei,SHE XiaoMan,YU Lin,LAN GuoBing,DENG MingGuang,HE ZiFu. Molecular Characteristic and Pathogenicity Analyses of Cucumber green mottle mosaic virus (CGMMV) Infecting Bottle Gourd in Lianzhou, Guangdong [J]. Scientia Agricultura Sinica, 2020, 53(5): 955-964. |
| [12] | LI YueYue,ZHOU WenPeng,LU SiQian,CHEN DeRong,DAI JianHong,GUO QiaoYou,LIU Yong,LI Fan,TAN GuanLin. Occurrence and Biological Characteristics of Tomato mottle mosaic virus on Solanaceae Crops in China [J]. Scientia Agricultura Sinica, 2020, 53(3): 539-550. |
| [13] | WANG BaoBao,GUO Cheng,SUN SuLi,XIA YuSheng,ZHU ZhenDong,DUAN CanXing. The Genetic Diversity, Pathogenicity, and Toxigenic Chemotypes of Fusarium graminearum Species Complex Causing Maize Ear Rot [J]. Scientia Agricultura Sinica, 2020, 53(23): 4777-4790. |
| [14] | SUN Qi,HE Fang,SHAO ShengNan,LIU Zheng,HUANG JiaFeng. Cloning and Functional Analysis of VdHP1 in Verticillium dahliae from Cotton [J]. Scientia Agricultura Sinica, 2020, 53(14): 2872-2884. |
| [15] | QI Yue,LÜ JunYuan,ZHANG Yue,WEI Jie,ZHANG Na,YANG WenXiang,LIU DaQun. Puccinia triticina Effector Protein Pt18906 Triggered Two-Layer Defense Reaction in TcLr27+31 [J]. Scientia Agricultura Sinica, 2020, 53(12): 2371-2384. |
|
||