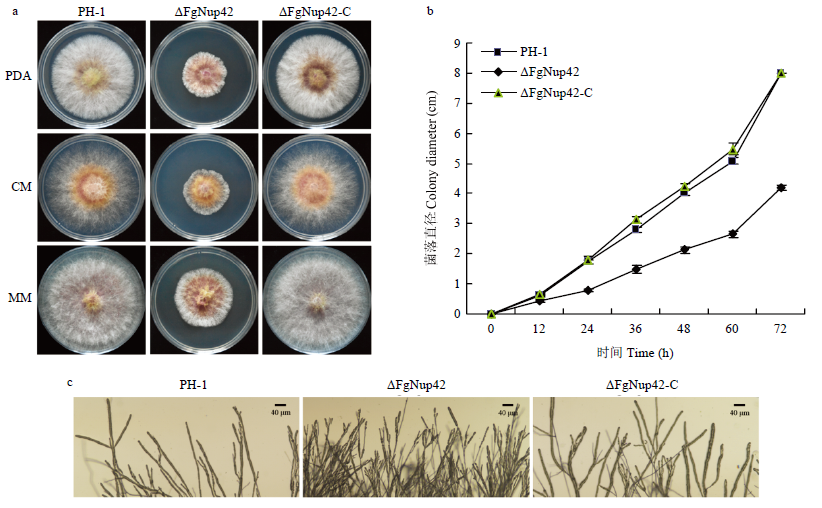
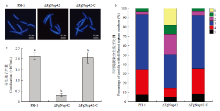
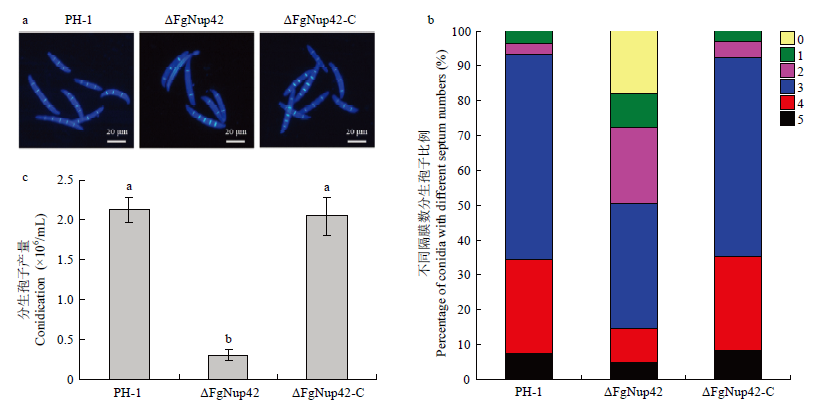
Scientia Agricultura Sinica ›› 2021, Vol. 54 ›› Issue (9): 1894-1903.doi: 10.3864/j.issn.0578-1752.2021.09.007
• PLANT PROTECTION • Previous Articles Next Articles
ZHANG ChengQi( ),LIAO LuLu,QI YongXia,DING KeJian,CHEN Li()
),LIAO LuLu,QI YongXia,DING KeJian,CHEN Li()
| [1] | 陈云, 王建强, 杨荣明, 马忠华. 小麦赤霉病发生危害形势及防控对策. 植物保护, 2017,43(5):11-17. |
| CHEN Y, WANG J Q, YANG R M, MA Z H. Current situation and management strategies of Fusarium head blight in China. Plant Protection, 2017,43(5):11-17. (in Chinese) | |
| [2] | MCMULLEN M, JONES R, GALLENBERG D. Scab of wheat and barley: A re-emerging disease of devastating impact. Plant Disease, 1997,81(12):1340-1348. |
| [3] | PESTKA J J, SMOLINSKI A T. Deoxynivalenol: Toxicology and potential effects on humans. Journal of Toxicology and Environmental Health, Part B Critical Reviews, 2005,8(1):39-69. |
| [4] | 史建荣, 刘馨, 仇剑波, 祭芳, 徐剑宏, 董飞, 殷宪超, 冉军舰. 小麦中镰刀菌毒素脱氧雪腐镰刀菌烯醇污染现状与防控研究进展. 中国农业科学, 2014,47(18):3641-3654. |
| SHI J R, LIU X, QIU J B, JI F, XU J H, DONG F, YIN X C, RAN J J. Deoxynivalenol contamination in wheat and its management. Scientia Agricultura Sinica, 2014,47(18):3641-3654. (in Chinese) | |
| [5] | CUOMO C A, GUELDENER U, XU J R, TRAIL F, TURGEON B G, DI PIETRO A, WALTON J D, MA L J, BAKER S E, REP M, et al. The Fusarium graminearum genome reveals a link between localized polymorphism and pathogen specialization. Science, 2007,317(5843):1400-1402. |
| [6] | SON H, SEO Y S, MIN K, PARK A R, LEE J, JIN J M, LIN Y, CAO P, HONG S Y, KIM E K, et al. A phenome-based functional analysis of transcription factors in the cereal head blight fungus, Fusarium graminearum. PLoS Pathogens, 2011,7(10):e1002310. |
| [7] | WANG C F, ZHANG S J, HOU R, ZHAO Z T, ZHENG Q, XU Q J, ZHENG D W, WANG G H, LIU H Q, GAO X L, MA J W, KISTLER H C, KANG Z S, XU J R. Functional analysis of the kinome of the wheat scab fungus Fusarium graminearum. PLoS Pathogens, 2011,7(12):e1002460. |
| [8] | YUN Y Z, LIU Z Y, YIN Y N, JIANG J H, CHEN Y, XU J R, MA Z H. Functional analysis of the Fusarium graminearum phosphatome. New Phytologist, 2015,207(1):119-134. |
| [9] | LIU H, WANG Q H, HE Y, CHEN L F, HAO C F, JIANG C, LI Y, DAI Y F, KANG Z S, XU J R. Genome-wide A-to-I RNA editing in fungi independent of ADAR enzymes. Genome Research, 2016,26(4):499-509. |
| [10] | CASEY A K, WENTE S R. Nuclear transport: Shifting gears in fungal nuclear and cytoplasmic organization. Current Biology, 2012,22(19):R846-R848. |
| [11] | RAICES M, D’ANGELO M A. Nuclear pore complexes and regulation of gene expression. Current Opinion in Cell Biology, 2017,46:26-32. |
| [12] | LIN D H, HOELZ A. The structure of the nuclear pore complex (An update). Annual Review of Biochemistry, 2019,88:725-783. |
| [13] | LIN D H, CORREIA A R, CAI S W, HUBER F M, JETTE C A, HOELZ A. Structural and functional analysis of mRNA export regulation by the nuclear pore complex. Nature Communications, 2018,9:2319. |
| [14] | MENON B B, SARMA N J, PASULA S, DEMINOFF S J, WILLIS K A, BARBARA K E, ANDREWS B, SANTANGELO G M. Reverse recruitment: The Nup84 nuclear pore subcomplex mediates Rap1/ Gcr1/Gcr2 transcriptional activation. Proceedings of the National Academy of Sciences of the United States of America, 2005,102(16):5749-5754. |
| [15] | CASOLARI J M, BROWN C R, KOMILI S, WEST J, HIERONYMUS H, SILVER P A. Genome-wide localization of the nuclear transport machinery couples transcriptional status and nuclear organization. Cell, 2004,117(4):427-439. |
| [16] | SCHMID M, ARIB G, LAEMMLI C, NISHIKAWA J, DURUSSEL T, LAEMMLI U K. Nup-PI: The nucleopore-promoter interaction of genes in yeast. Molecular Cell, 2006,21(3):379-391. |
| [17] | STUTZ F, NEVILLE M, ROSBASH M. Identification of a novel nuclear pore-associated protein as a functional target of the HIV-1 Rev protein in yeast. Cell, 1995,82(3):495-506. |
| [18] | MURPHY R, WENTE S R. An RNA-export mediator with an essential nuclear export signal. Nature, 1996,383(6598):357-360. |
| [19] | BOLGER T A, FOLKMANN A W, TRAN E J, WENTE S R. The mRNA export factor Gle1 and inositol hexakisphosphate regulate distinct stages of translation. Cell, 2008,134(4):624-633. |
| [20] | GROSS T, SIEPMANN A, STURM D, WINDGASSEN M, SCARCELLI J J, SEEDORF M, COLE C N, KREBBER H. The DEAD-box RNA helicase Dbp5 functions in translation termination. Science, 2007,315(5812):646-649. |
| [21] | ADAMS R L, TERRY L J, WENTE S R. Nucleoporin FG domains facilitate mRNP remodeling at the cytoplasmic face of the nuclear pore complex. Genetics, 2014,197(4):1213-1224. |
| [22] | STRASSER K, BASSLER J, HURT E. Binding of the Mex67p/Mtr2p heterodimer to FXFG, GLFG, and FG repeat nucleoporins is essential for nuclear mRNA export. The Journal of Cell Biology, 2000,150(4):695-706. |
| [23] | KENDIRGI F, REXER D J, ALCAZAR-ROMAN A R, ONISHKO H M, WENTE S R. Interaction between the shuttling mRNA export factor Gle1 and the nucleoporin hCG1: A conserved mechanism in the export of Hsp70 mRNA. Molecular Biology of the Cell, 2005,16(9):4304-4315. |
| [24] | SAAVEDRA C A, HAMMELL C M, HEATH C V, COLE C N. Yeast heat shock mRNAs are exported through a distinct pathway defined by Rip1p. Genes and Development, 1997,11(21):2845-2856. |
| [25] | MILLER A L, SUNTHARALINGAM M, JOHNSON S L, AUDHYA A, EMR S D, WENTE S R. Cytoplasmic inositol hexakisphosphate production is sufficient for mediating the Gle1-mRNA export pathway. The Journal of Biological Chemistry, 2004,279(49):51022-51032. |
| [26] | STUTZ F, KANTOR J, ZHANG D, MCCARTHY T, NEVILLE M, ROSBASH M. The yeast nucleoporin Rip1p contributes to multiple export pathways with no essential role for its FG-repeat region. Genes and Development, 1997,11(21):2857-2868. |
| [27] | NICHOLSON P. The Fusarium laboratory manual. Plant Pathology, 2007,56(6):1037. |
| [28] | YU J H, HAMARI Z, HAN K H, SEO J A, REYES-DOMINGUEZ Y, SCAZZOCCHIO C. Double-joint PCR: A PCR-based molecular tool for gene manipulations in filamentous fungi. Fungal Genetics and Biology, 2004,41(11):973-981. |
| [29] | PROCTOR R H, HOHN T M, MCCORMICK S P. Reduced virulence of Gibberella zeae caused by disruption of a trichothecene toxin biosynthetic gene. Molecular Plant-Microbe Interactions, 1995,8(4):593-601. |
| [30] | BRUNO K S, TENJO F, LI L, HAMER J E, XU J R. Cellular localization and role of kinase activity of PMK1 in Magnaporthe grisea. Eukaryotic Cell, 2004,3(6):1525-1532. |
| [31] | CHEN Y, ZHENG S Y, JU Z, ZHANG C Q, TANG G F, WANG J, WEN Z Y, CHEN W, MA Z H. Contribution of peroxisomal docking machinery to mycotoxin biosynthesis, pathogenicity and pexophagy in the plant pathogenic fungus Fusarium graminearum. Environmental Microbiology, 2018,20(9):3224-3245. |
| [32] | JI F, XU J H, LIU X, YIN X C, SHI J R. Natural occurrence of deoxynivalenol and zearalenone in wheat from Jiangsu Province, China. Food Chemistry, 2014,157:393-397. |
| [33] | MENKE J, DONG Y H, KISTLER H C. Fusarium graminearum Tri12p influences virulence to wheat and trichothecene accumulation. Molecular Plant-Microbe Interactions, 2012,25(11):1408-1418. |
| [34] | TRAIL F. For blighted waves of grain: Fusarium graminearum in the postgenomics era. Plant Physiology, 2009,149(1):103-110. |
| [35] | SEONG K Y, PASQUALI M, ZHOU X, SONG J, HILBURN K, MCCORMICK S, DONG Y, XU J R, KISTLER H C. Global gene regulation by Fusarium transcription factors Tri6 and Tri10 reveals adaptations for toxin biosynthesis. Molecular Microbiology, 2009,72(2):354-367. |
| [36] | FABRE E, HURT E. Yeast genetics to dissect the nuclear pore complex and nucleocytoplasmic trafficking. Annual Review of Genetics, 1997,31:277-313. |
| [37] | ADAMS R L, MASON A C, GLASS L, ADITI , WENTE S R. Nup42 and IP6 coordinate Gle1 stimulation of Dbp5/DDX19B for mRNA export in yeast and human cells. Traffic, 2017,18(12):776-790. |
| [38] | ZHENG D W, ZHANG S J, ZHOU X Y, WANG C F, XIANG P, ZHENG Q, XU J R. The FgHOG1 pathway regulates hyphal growth, stress responses, and plant infection in Fusarium graminearum. PLoS ONE, 2012,7(11):e49495. |
| [39] | YUN Y Z, LIU Z Y, ZHANG J Z, SHIM W B, CHEN Y, MA Z H. The MAPKK FgMkk1 of Fusarium graminearum regulates vegetative differentiation, multiple stress response, and virulence via the cell wall integrity and high-osmolarity glycerol signaling pathways. Environmental Microbiology, 2014,16(7):2023-2037. |
| [40] | LIU Z Y, JIAN Y Q, CHEN Y, KISTLER H C, HE P, MA Z H, YIN Y N. A phosphorylated transcription factor regulates sterol biosynthesis in Fusarium graminearum. Nature Communications, 2019,10:1228. |
| [41] | LIU N, WU S Q, DAWOOD D H, TANG G F, ZHANG C Q, LIANG J T, CHEN Y, MA Z H. The b-ZIP transcription factor FgTfmI is required for the fungicide phenamacril tolerance and pathogenecity in Fusarium graminearum. Pest Management Science, 2019,75(12):3312-3322. |
| [42] | BAI G H, DESJARDINS A E, PLATTNER R D. Deoxynivalenol- nonproducing Fusarium graminearum causes initial infection, but does not cause disease spread in wheat spikes. Mycopathologia, 2002,153(2):91-98. |
| [1] | TAO Bu, QI YongZhi, QU Yun, CAO ZhiYan, ZHAO XuSheng, ZHEN WenChao. Construction and Verification of Fusarium Head Blight Prediction Model in Haihe Plain Based on Boosted Regression Tree [J]. Scientia Agricultura Sinica, 2021, 54(18): 3860-3870. |
| [2] | ZHANG ChengQi,WANG XiaoYan,CHEN Li. The Actin Binding Protein FgAbp1 is Involved in Growth, Development and Toxisome Formation in Fusarium graminearum [J]. Scientia Agricultura Sinica, 2021, 54(13): 2759-2768. |
| [3] | WANG BaoBao,GUO Cheng,SUN SuLi,XIA YuSheng,ZHU ZhenDong,DUAN CanXing. The Genetic Diversity, Pathogenicity, and Toxigenic Chemotypes of Fusarium graminearum Species Complex Causing Maize Ear Rot [J]. Scientia Agricultura Sinica, 2020, 53(23): 4777-4790. |
| [4] | SUN Qi,HE Fang,SHAO ShengNan,LIU Zheng,HUANG JiaFeng. Cloning and Functional Analysis of VdHP1 in Verticillium dahliae from Cotton [J]. Scientia Agricultura Sinica, 2020, 53(14): 2872-2884. |
|
||