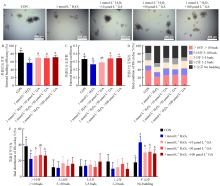
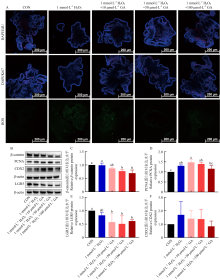
Scientia Agricultura Sinica ›› 2025, Vol. 58 ›› Issue (22): 4771-4785.doi: 10.3864/j.issn.0578-1752.2025.22.016
• ANIMAL SCIENCE·VETERINARY SCIENCE • Previous Articles Next Articles
LI RuiTong( ), CHEN QingMei, XU JianFang, ZHANG JunMin, SI Wei(), ZHANG TieYing()
), CHEN QingMei, XU JianFang, ZHANG JunMin, SI Wei(), ZHANG TieYing()
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [1] | WANG Wei, LUO ChunHai, JIA HongDou, LIU JiaJin, LI DanYang, FU ShiXin. Effect of Gln on Endoplasmic Reticulum Stress in Retained Fetal Membranes Cows Under Oxidative Stress via the PI3K/AKT Pathway [J]. Scientia Agricultura Sinica, 2025, 58(7): 1451-1462. |
| [2] | LI XiangYu, LIU JianZhuo, HU DanDan, LIU GengYu, CHEN LiangYu, LI Bing, DU WanLi, SONG Bo. Characterization of Maize Germplasm Resistance to Common Smut and Analysis of Physiological Differences [J]. Scientia Agricultura Sinica, 2025, 58(13): 2504-2521. |
| [3] | ZHANG Bing, YANG YanYan, FENG QianHui, SHI Wen, FANG YiZhen, HUANG JiaBao, SHI DeShun. Effects of Sodium Selenite on the in vitro Maturation of Porcine Oocytes and Their Embryonic Development Potentials [J]. Scientia Agricultura Sinica, 2024, 57(17): 3482-3493. |
| [4] | CUI HongJie, LU ChunTing, PAN LiQin, HU Hui, ZHONG PeiYun, ZHU JieYing, ZHANG KaiZhao, HUANG XiaoHong. Curcumin Alleviates Zearalenone-Induced Oxidative Damage in Porcine Renal Epithelial Cells via SIRT1/FOXO1 Pathway [J]. Scientia Agricultura Sinica, 2023, 56(5): 1007-1018. |
| [5] | XI MengXue, SHEN Dan, SHI YiFan, LI ChunMei. Effects of TBHQ on Pyroptosis, Necroptosis and Inflammatory Damage of Chicken Embryonic Lung Tissues Induced by PM2.5 from Chicken Houses [J]. Scientia Agricultura Sinica, 2023, 56(4): 779-787. |
| [6] | TAO WenJing, ZHANG ZiTing, LIU Yuan, SONG Dan, LI XiangChen. Inhibitory Effect of N-acetylcysteine on Bisphenol A-Induced Apoptosis and Inflammatory Response in Porcine Kidney Cells [J]. Scientia Agricultura Sinica, 2023, 56(3): 549-558. |
| [7] | ZHANG Nan, HAN GuangJie, LIU Qin, LI ChuanMing, QI JianHang, LU YuRong, XIA Yang, XU Jian. Response and Function of the Transcription Factor CncC in Cnaphalocrocis medinalis Infected with Baculovirus CnmeGV [J]. Scientia Agricultura Sinica, 2023, 56(13): 2491-2503. |
| [8] | YANG ChangPei,WANG NaiXiu,WANG Kai,HUANG ZiQing,LIN HaiLan,ZHANG Li,ZHANG Chen,FENG LuQiu,GAN Ling. Effects and Mechanisms of Exogenous GABA Against Oxidative Stress in Piglets [J]. Scientia Agricultura Sinica, 2022, 55(17): 3437-3449. |
| [9] | JIANG ChunHui,SUN XuDong,TANG Yan,LUO ShengBin,XU Chuang,CHEN YuanYuan. Curcumin Alleviates H2O2-Induced Oxidative Stress in Bovine Mammary Epithelial Cells Via the Nrf2 Signaling Pathway [J]. Scientia Agricultura Sinica, 2021, 54(8): 1787-1794. |
| [10] | LÜ ChuYang,DENG PingChuan,ZHANG XiaoLi,SUN YuChao,LIANG WuSheng,HU DongWei. Transcriptomic Analysis of Sclerotia Formation Induced by Low Temperature in Villosiclava virens [J]. Scientia Agricultura Sinica, 2020, 53(22): 4571-4583. |
| [11] | LI HanTong,JIA ChengLi,ZHANG ShuWen,LU Jing,PANG XiaoYang,LIU Lu,LÜ JiaPing. Chromium (III) Stress Alleviation by Sulfur Compounds During Chromium Bio-enrichment by Saccharomyces cerevisiae [J]. Scientia Agricultura Sinica, 2019, 52(6): 1078-1089. |
| [12] | DU Jiao, WANG YaBo, LI XueHua, HUANG ZhiQiang, YANG YuHeng, BI ChaoWei, YU Yang. Function analysis ofγ-glutamyl phosphate reductase-encoded gene SsGPR1 in Sclerotinia sclerotiorum [J]. Scientia Agricultura Sinica, 2018, 51(19): 3694-3703. |
| [13] | TANG Qin, DENG Yuan-yuan, ZHANG Rui-fen, ZHANG Yan, ZHANG Ming-wei, WEI Zhen-cheng, TANG Xiao-jun, LIU Lei, TI Hui-hui, MA Yong-xuan. Effect of Momordica charantia Fruit Aqueous Extract on Serum Lipids Metabolic Disorder in Restraint Mice [J]. Scientia Agricultura Sinica, 2014, 47(16): 3300-3307. |
| [14] | ZHAO Jiao, ZHOU Zhao-Hong, LIANG Xiao-Fang, MAO Xiang-Bing, CHEN Dai-Wen, YU Bing. Effects of GSPs and VE on Growth Performance, Serum Redox Status and Hepatic Oxidative Damage in Piglets Under Oxidative Stress [J]. Scientia Agricultura Sinica, 2013, 46(19): 4157-4164. |
| [15] | SONG Gui-Fang-., FAN Wei-Li, WANG Jun-Juan, WANG De-Long, WANG Shuai, ZHOU Kai, YE Wu-Wei. Cloning and Characterization of Drought-stress Responsive Gene GhGR in Gossypium hirsutum L. [J]. Scientia Agricultura Sinica, 2012, 45(8): 1644-1652. |
|
||