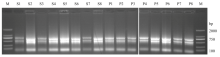
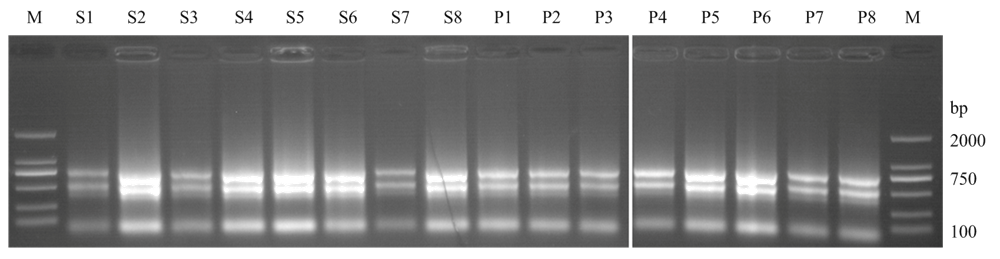
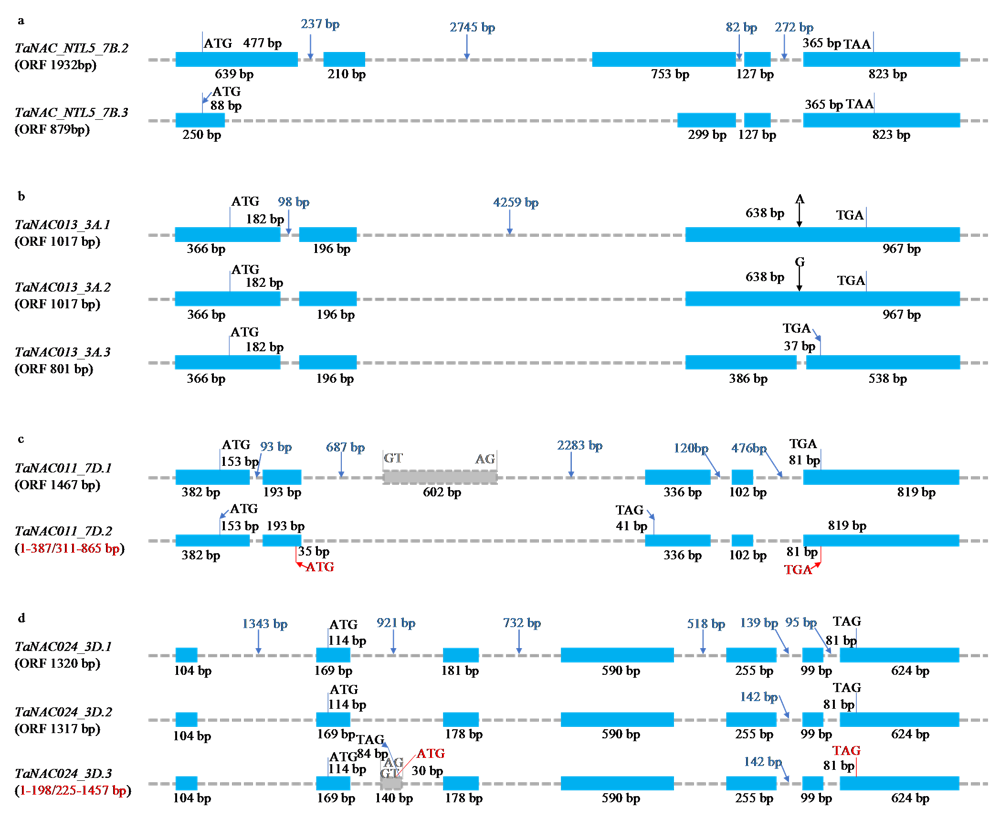
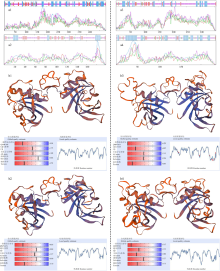
Scientia Agricultura Sinica ›› 2021, Vol. 54 ›› Issue (22): 4709-4727.doi: 10.3864/j.issn.0578-1752.2021.22.001
• CROP GENETICS & BREEDING·GERMPLASM RESOURCES·MOLECULAR GENETICS • Previous Articles Next Articles
LÜ ShiKai( ),MA XiaoLong,ZHANG Min,DENG PingChuan,CHEN ChunHuan,ZHANG Hong(),LIU XinLun(),JI WanQuan()
),MA XiaoLong,ZHANG Min,DENG PingChuan,CHEN ChunHuan,ZHANG Hong(),LIU XinLun(),JI WanQuan()
| [1] |
FUJITA M, FUJITA Y, MARUYAMA K, SEKI M, HIRATSU K, OHME-TAKAGI M, TRAN L S, YAMAGUCHI-SHINOZAKI K, SHINOZAKI K. A dehydration-induced NAC protein, RD26, is involved in a novel ABA-dependent stress-signaling pathway. The Plant Journal, 2004, 39(6): 863-876.
doi: 10.1111/tpj.2004.39.issue-6 |
| [2] |
TRAN L S, NAKASHIMA K, SAKUMA Y, SIMPSON S D, FUJITA Y, MARUYAMA K, FUJITA M, SEKI M, SHINOZAKI K, YAMAGUCHI-SHINOZAKI K. Isolation and functional analysis of Arabidopsis stress-inducible NAC transcription factors that bind to a drought-responsive cis-element in the early responsive to dehydration stress 1 promoter. The Plant Cell, 2004, 16(9): 2481-2498.
doi: 10.1105/tpc.104.022699 |
| [3] |
GUO Y, GAN S. AtNAP, a NAC family transcription factor, has an important role in leaf senescence. The Plant Journal, 2006, 46(4): 601-612.
doi: 10.1111/tpj.2006.46.issue-4 |
| [4] | HU H, DAI M, YAO J, XIAO B, LI X, ZHANG Q, XIONG L. Overexpressing a NAM, ATAF, and CUC (NAC) transcription factor enhances drought resistance and salt tolerance in rice. Proceedings of the National Academy of Sciences of the United States of America, 2006, 103(35): 12987-12992. |
| [5] |
XU B, OHTANI M, YAMAGUCHI M, TOYOOKA K, WAKAZAKI M, SATO M, KUBO M, NAKANO Y, SANO R, HIWATASHI Y, MURATA T, KURATA T, YONEDA A, KATO K, HASEBE M, DEMURA T. Contribution of NAC transcription factors to plant adaptation to land. Science, 2014, 343(6178): 1505-1508.
doi: 10.1126/science.1248417 |
| [6] |
LV S, GUO H, ZHANG M, WANG Q, ZHANG H, JI W. Large-scale cloning and comparative analysis of TaNAC genes in response to stripe rust and powdery mildew in wheat (Triticum aestivum L.). Genes (Basel), 2020, 11(9): 1073.
doi: 10.3390/genes11091073 |
| [7] |
FENG H, DUAN X, ZHANG Q, LI X, WANG B, HUANG L, WANG X, KANG Z. The target gene of tae-miR164, a novel NAC transcription factor from the NAM subfamily, negatively regulates resistance of wheat to stripe rust. Molecular Plant Pathology, 2014, 15(3): 284-296.
doi: 10.1111/mpp.2014.15.issue-3 |
| [8] |
RIECHMANN J L, HEARD J, MARTIN G, REUBER L, JIANG C, KEDDIE J, ADAM L, PINEDA O, RATCLIFFE O J, SAMAHA R R, CREELMAN R, PILGRIM M, BROUN P, ZHANG J Z, GHANDEHARI D, SHERMAN B K, YU G. Arabidopsis transcription factors: Genome-wide comparative analysis among eukaryotes. Science, 2000, 290(5499): 2105-2110.
doi: 10.1126/science.290.5499.2105 |
| [9] |
SOUER E, VANHOUWELINGEN A, KLOOS D, MOL J, KOES R. The no apical meristem gene of petunia is required for pattern formation in embryos and flowers and is expressed at meristem and primordia boundaries. Cell, 1996, 85(2): 159-170.
doi: 10.1016/S0092-8674(00)81093-4 |
| [10] |
AIDA M, ISHIDA T, FUKAKI H, FUJISAWA H, TASAKA M. Genes involved in organ separation in Arabidopsis: An analysis of the cup-shaped cotyledon mutant. The Plant Cell, 1997, 9(6): 841-857.
doi: 10.1105/tpc.9.6.841 |
| [11] |
OOKA H, SATOH K, DOI K, NAGATA T, OTOMO Y, MURAKAMI K, MATSUBARA K, OSATO N, KAWAI J, CARNINCI P, HAYASHIZAKI Y, SUZUKI K, KOJIMA K, TAKAHARA Y, YAMAMOTO K, KIKUCHI S. Comprehensive analysis of NAC family genes in Oryza sativa and Arabidopsis thaliana. DNA Research, 2003, 10(6): 239-247.
doi: 10.1093/dnares/10.6.239 |
| [12] |
OLSEN A N, ERNST H A, LEGGIO L L, SKRIVER K. NAC transcription factors: Structurally distinct, functionally diverse. Trends in Plant Science, 2005, 10(2): 79-87.
doi: 10.1016/j.tplants.2004.12.010 |
| [13] |
MUROZUKA E, MASSANGE-SANCHEZ J A, NIELSEN K, GREGERSEN P L, BRAUMANN I. Genome wide characterization of barley NAC transcription factors enables the identification of grain-specific transcription factors exclusive for the Poaceae family of monocotyledonous plants. PLoS ONE, 2018, 13(12): e0209769.
doi: 10.1371/journal.pone.0209769 |
| [14] | TIAN F, YANG D C, MENG Y Q, JIN J, GAO G. PlantRegMap: Charting functional regulatory maps in plants. Nucleic Acids Research, 2020, 48(D1): D1104-D1113. |
| [15] |
GUERIN C, ROCHE J, ALLARD V, RAVEL C, MOUZEYAR S, BOUZIDI M F. Genome-wide analysis, expansion and expression of the NAC family under drought and heat stresses in bread wheat (T. aestivum L.). PLoS ONE, 2019, 14(3): e0213390.
doi: 10.1371/journal.pone.0213390 |
| [16] |
ZHAO D, DERKX A P, LIU D C, BUCHNER P, HAWKESFORD M J. Overexpression of a NAC transcription factor delays leaf senescence and increases grain nitrogen concentration in wheat. Plant Biology (Stuttgart), 2015, 17(4): 904-913.
doi: 10.1111/plb.2015.17.issue-4 |
| [17] |
HARRINGTON S A, OVEREND L E, COBO N, BORRILL P, UAUY C. Conserved residues in the wheat (Triticum aestivum) NAM-A1 NAC domain are required for protein binding and when mutated lead to delayed peduncle and flag leaf senescence. BMC Plant Biology, 2019, 19(1): 407.
doi: 10.1186/s12870-019-2022-5 |
| [18] |
XUE G P, BOWER N I, MCINTYRE C L, RIDING G A, KAZAN K, SHORTER R. TaNAC69 from the NAC superfamily of transcription factors is up-regulated by abiotic stresses in wheat and recognises two consensus DNA-binding sequences. Functional Plant Biology, 2006, 33(1): 43-57.
doi: 10.1071/FP05161 |
| [19] | ZHANG L, ZHANG L, XIA C, ZHAO G, JIA J, KONG X. The novel wheat transcription factor TaNAC47 enhances multiple abiotic stress tolerances in transgenic plants. Front Plant Science, 2015, 6: 1174. |
| [20] |
PEROCHON A, KAHLA A, VRANIC M, JIA J, MALLA K B, CRAZE M, WALLINGTON E, DOOHAN F M. A wheat NAC interacts with an orphan protein and enhances resistance to Fusarium head blight disease. Plant Biotechnology Journal, 2019, 17(10): 1892-1904.
doi: 10.1111/pbi.v17.10 |
| [21] |
WANG B, WEI J, SONG N, WANG N, ZHAO J, KANG Z. A novel wheat NAC transcription factor, TaNAC30, negatively regulates resistance of wheat to stripe rust. Journal of Integrative Plant Biology, 2018, 60(5): 432-443.
doi: 10.1111/jipb.v60.5 |
| [22] |
PURANIK S, SAHU P P, SRIVASTAVA P S, PRASAD M. NAC proteins: Regulation and role in stress tolerance. Trends in Plant Science, 2012, 17(6): 369-381.
doi: 10.1016/j.tplants.2012.02.004 |
| [23] |
AMERES S L, ZAMORE P D. Diversifying microRNA sequence and function. Nature Reviews. Molecular Cell Biology, 2013, 14(8): 475-488.
doi: 10.1038/nrm3611 |
| [24] |
ZHANG H, MAO R, WANG Y, ZHANG L, WANG C, LV S, LIU X, WANG Y, JI W. Transcriptome-wide alternative splicing modulation during plant-pathogen interactions in wheat. Plant Science, 2019, 288: 110160.
doi: 10.1016/j.plantsci.2019.05.023 |
| [25] |
ZHANG X M, ZHANG Q, PEI C L, LI X, HUANG X L, CHANG C Y, WANG X J, HUANG L L, KANG Z S. TaNAC2 is a negative regulator in the wheat-stripe rust fungus interaction at the early stage. Physiological and Molecular Plant Pathology, 2018, 102: 144-153.
doi: 10.1016/j.pmpp.2018.02.002 |
| [26] |
WANG H-L, ZHANG Y, WANG T, YANG Q, YANG Y, LI Z, LI B, WEN X, LI W, YIN W, XIA X, GUO H, LI Z. An alternative splicing variant of PtRD26 delays leaf senescence by regulating multiple NAC transcription factors in Populus. The Plant Cell, 2021, 33: 1594-1614.
doi: 10.1093/plcell/koab046 |
| [27] |
REDDY A S, MARQUEZ Y, KALYNA M, BARTA A. Complexity of the alternative splicing landscape in plants. The Plant Cell, 2013, 25(10): 3657-3683.
doi: 10.1105/tpc.113.117523 |
| [28] |
XUE F, JI W, WANG C, ZHANG H, YANG B. High-density mapping and marker development for the powdery mildew resistance gene PmAS846 derived from wild emmer wheat (Triticum turgidum var. dicoccoides). Theoretical and Applied Genetics, 2012, 124(8): 1549-1560.
doi: 10.1007/s00122-012-1809-7 |
| [29] |
ZHANG H, YANG Y, WANG C, LIU M, LI H, FU Y, WANG Y, NIE Y, LIU X, JI W. Large-scale transcriptome comparison reveals distinct gene activations in wheat responding to stripe rust and powdery mildew. BMC Genomics, 2014, 15(1): 898.
doi: 10.1186/1471-2164-15-898 |
| [30] |
ZHANG H, MAO R, WANG Y, ZHANG L, WANG C, LV S, LIU X, WANG Y, JI W. Transcriptome-wide alternative splicing modulation during plant-pathogen interactions in wheat. Plant Science, 2019, 288: 110160.
doi: 10.1016/j.plantsci.2019.05.023 |
| [31] |
SANCHEZ-MARTIN J, WIDRIG V, HERREN G, WICKER T, ZBINDEN H, GRONNIER J, SPORRI L, PRAZ C R, HEUBERGER M, KOLODZIEJ M C, ISAKSSON J, STEUERNAGEL B, KARAFIATOVA M, DOLEZEL J, ZIPFEL C, KELLER B. Wheat Pm4 resistance to powdery mildew is controlled by alternative splice variants encoding chimeric proteins. Nature Plants, 2021, 7(3): 327-341.
doi: 10.1038/s41477-021-00869-2 |
| [32] |
GAO P, QUILICHINI T D, ZHAI C, QIN L, NILSEN K T, LI Q, SHARPE A G, KOCHIAN L V, ZOU J, REDDY A S N, WEI Y, POZNIAK C, PATTERSON N, GILLMOR C S, DATLA R, XIANG D. Alternative splicing dynamics and evolutionary divergence during embryogenesis in wheat species. Plant Biotechnology Journal, 2021, 19: 1624-1643.
doi: 10.1111/pbi.v19.8 |
| [33] |
KOZOMARA A, BIRGAOANU M, GRIFFITHS-JONES S. miRBase: From microRNA sequences to function. Nucleic Acids Research, 2019, 47(D1): D155-D162.
doi: 10.1093/nar/gky1141 |
| [34] |
FANG Y, XIE K, XIONG L. Conserved miR164-targeted NAC genes negatively regulate drought resistance in rice. Journal of Experimental Botany, 2014, 65(8): 2119-2135.
doi: 10.1093/jxb/eru072 |
| [35] |
KIM J H, WOO H R, KIM J, LIM P O, LEE I C, CHOI S H, HWANG D, NAM H G. Trifurcate feed-forward regulation of age-dependent cell death involving miR164 in Arabidopsis. Science, 2009, 323(5917): 1053-1057.
doi: 10.1126/science.1166386 |
| [1] | CHEN JiHao, ZHOU JieGuang, QU XiangRu, WANG SuRong, TANG HuaPing, JIANG Yun, TANG LiWei, $\boxed{\hbox{LAN XiuJin}}$, WEI YuMing, ZHOU JingZhong, MA Jian. Mapping and Analysis of QTL for Embryo Size-Related Traits in Tetraploid Wheat [J]. Scientia Agricultura Sinica, 2023, 56(2): 203-216. |
| [2] | YAN YanGe, ZHANG ShuiQin, LI YanTing, ZHAO BingQiang, YUAN Liang. Effects of Dextran Modified Urea on Winter Wheat Yield and Fate of Nitrogen Fertilizer [J]. Scientia Agricultura Sinica, 2023, 56(2): 287-299. |
| [3] | XU JiuKai, YUAN Liang, WEN YanChen, ZHANG ShuiQin, LI YanTing, LI HaiYan, ZHAO BingQiang. Nitrogen Fertilizer Replacement Value of Livestock Manure in the Winter Wheat Growing Season [J]. Scientia Agricultura Sinica, 2023, 56(2): 300-313. |
| [4] | ZHAO HaiXia,XIAO Xin,DONG QiXin,WU HuaLa,LI ChengLei,WU Qi. Optimization of Callus Genetic Transformation System and Its Application in FtCHS1 Overexpression in Tartary Buckwheat [J]. Scientia Agricultura Sinica, 2022, 55(9): 1723-1734. |
| [5] | WANG HaoLin,MA Yue,LI YongHua,LI Chao,ZHAO MingQin,YUAN AiJing,QIU WeiHong,HE Gang,SHI Mei,WANG ZhaoHui. Optimal Management of Phosphorus Fertilization Based on the Yield and Grain Manganese Concentration of Wheat [J]. Scientia Agricultura Sinica, 2022, 55(9): 1800-1810. |
| [6] | TANG HuaPing,CHEN HuangXin,LI Cong,GOU LuLu,TAN Cui,MU Yang,TANG LiWei,LAN XiuJin,WEI YuMing,MA Jian. Unconditional and Conditional QTL Analysis of Wheat Spike Length in Common Wheat Based on 55K SNP Array [J]. Scientia Agricultura Sinica, 2022, 55(8): 1492-1502. |
| [7] | MA XiaoYan,YANG Yu,HUANG DongLin,WANG ZhaoHui,GAO YaJun,LI YongGang,LÜ Hui. Annual Nutrients Balance and Economic Return Analysis of Wheat with Fertilizers Reduction and Different Rotations [J]. Scientia Agricultura Sinica, 2022, 55(8): 1589-1603. |
| [8] | LIU Shuo,ZHANG Hui,GAO ZhiYuan,XU JiLi,TIAN Hui. Genetic Variations of Potassium Harvest Index in 437 Wheat Varieties [J]. Scientia Agricultura Sinica, 2022, 55(7): 1284-1300. |
| [9] | WANG YangYang,LIU WanDai,HE Li,REN DeChao,DUAN JianZhao,HU Xin,GUO TianCai,WANG YongHua,FENG Wei. Evaluation of Low Temperature Freezing Injury in Winter Wheat and Difference Analysis of Water Effect Based on Multivariate Statistical Analysis [J]. Scientia Agricultura Sinica, 2022, 55(7): 1301-1318. |
| [10] | GOU ZhiWen,YIN Wen,CHAI Qiang,FAN ZhiLong,HU FaLong,ZHAO Cai,YU AiZhong,FAN Hong. Analysis of Sustainability of Multiple Cropping Green Manure in Wheat-Maize Intercropping After Wheat Harvested in Arid Irrigation Areas [J]. Scientia Agricultura Sinica, 2022, 55(7): 1319-1331. |
| [11] | ZHI Lei,ZHE Li,SUN NanNan,YANG Yang,Dauren Serikbay,JIA HanZhong,HU YinGang,CHEN Liang. Genome-Wide Association Analysis of Lead Tolerance in Wheat at Seedling Stage [J]. Scientia Agricultura Sinica, 2022, 55(6): 1064-1081. |
| [12] | QIN YuQing,CHENG HongBo,CHAI YuWei,MA JianTao,LI Rui,LI YaWei,CHANG Lei,CHAI ShouXi. Increasing Effects of Wheat Yield Under Mulching Cultivation in Northern of China: A Meta-Analysis [J]. Scientia Agricultura Sinica, 2022, 55(6): 1095-1109. |
| [13] | CAI WeiDi,ZHANG Yu,LIU HaiYan,ZHENG HengBiao,CHENG Tao,TIAN YongChao,ZHU Yan,CAO WeiXing,YAO Xia. Early Detection on Wheat Canopy Powdery Mildew with Hyperspectral Imaging [J]. Scientia Agricultura Sinica, 2022, 55(6): 1110-1126. |
| [14] | ZONG Cheng, WU JinXin, ZHU JiuGang, DONG ZhiHao, LI JunFeng, SHAO Tao, LIU QinHua. Effects of Additives on the Fermentation Quality of Agricultural By-Products and Wheat Straw Mixed Silage [J]. Scientia Agricultura Sinica, 2022, 55(5): 1037-1046. |
| [15] | MA HongXiang, WANG YongGang, GAO YuJiao, HE Yi, JIANG Peng, WU Lei, ZHANG Xu. Review and Prospect on the Breeding for the Resistance to Fusarium Head Blight in Wheat [J]. Scientia Agricultura Sinica, 2022, 55(5): 837-855. |
|
||