Scientia Agricultura Sinica ›› 2023, Vol. 56 ›› Issue (9): 1633-1645.doi: 10.3864/j.issn.0578-1752.2023.09.002
• CROP GENETICS & BREEDING·GERMPLASM RESOURCES·MOLECULAR GENETICS • Previous Articles Next Articles
PENG HaiXia1( ), KA DeYan2, ZHANG TianXing3, ZHOU MengDie2, WU LinNan2, XIN ZhuanXia1, ZHAO HuiXian2, MA Meng2()
), KA DeYan2, ZHANG TianXing3, ZHOU MengDie2, WU LinNan2, XIN ZhuanXia1, ZHAO HuiXian2, MA Meng2()
| [1] |
doi: 10.1111/nph.2009.185.issue-1 |
| [2] |
doi: 10.1093/oxfordjournals.aob.a087220 |
| [3] |
doi: 10.1016/j.pbi.2013.11.005 pmid: 24507492 |
| [4] |
doi: 10.1016/j.cub.2012.02.010 |
| [5] |
doi: 10.1093/jxb/ert025 pmid: 23404902 |
| [6] |
doi: 10.1016/j.cub.2005.12.026 pmid: 16461280 |
| [7] |
doi: 10.1101/gad.463608 |
| [8] |
doi: 10.1105/tpc.113.115063 |
| [9] |
doi: 10.1038/sj.emboj.7601270 |
| [10] |
doi: 10.1111/tpj.2009.60.issue-6 |
| [11] |
|
| [12] |
doi: 10.1105/tpc.107.055467 |
| [13] |
|
| [14] |
doi: 10.1186/1471-2164-6-92 |
| [15] |
doi: 10.1046/j.1365-313X.1999.00523.x |
| [16] |
pmid: 18061566 |
| [17] |
doi: 10.3390/ijms23084219 |
| [18] |
|
| [19] |
doi: 10.1105/tpc.108.058180 |
| [20] |
doi: 10.1371/journal.pgen.1009043 |
| [21] |
doi: 10.1104/pp.113.218214 |
| [22] |
doi: 10.1105/tpc.12.9.1541 |
| [23] |
doi: 10.1111/tpj.2012.70.issue-6 |
| [24] |
|
| [25] |
|
| [26] |
doi: 10.1016/S1672-6308(13)60140-7 |
| [27] |
doi: 10.1111/tpj.12223 pmid: 23621326 |
| [28] |
|
| [29] |
doi: 10.1093/mp/sst107 pmid: 23775595 |
| [30] |
doi: 10.1111/tpj.12896 |
| [31] |
doi: 10.1111/pbi.v20.1 |
| [32] |
doi: 10.1038/ncomms14752 pmid: 28300078 |
| [33] |
doi: 10.1093/plphys/kiaa034 pmid: 33721894 |
| [34] |
doi: 10.1016/j.cub.2010.01.039 |
| [35] |
doi: 10.1093/plphys/kiab067 |
| [36] |
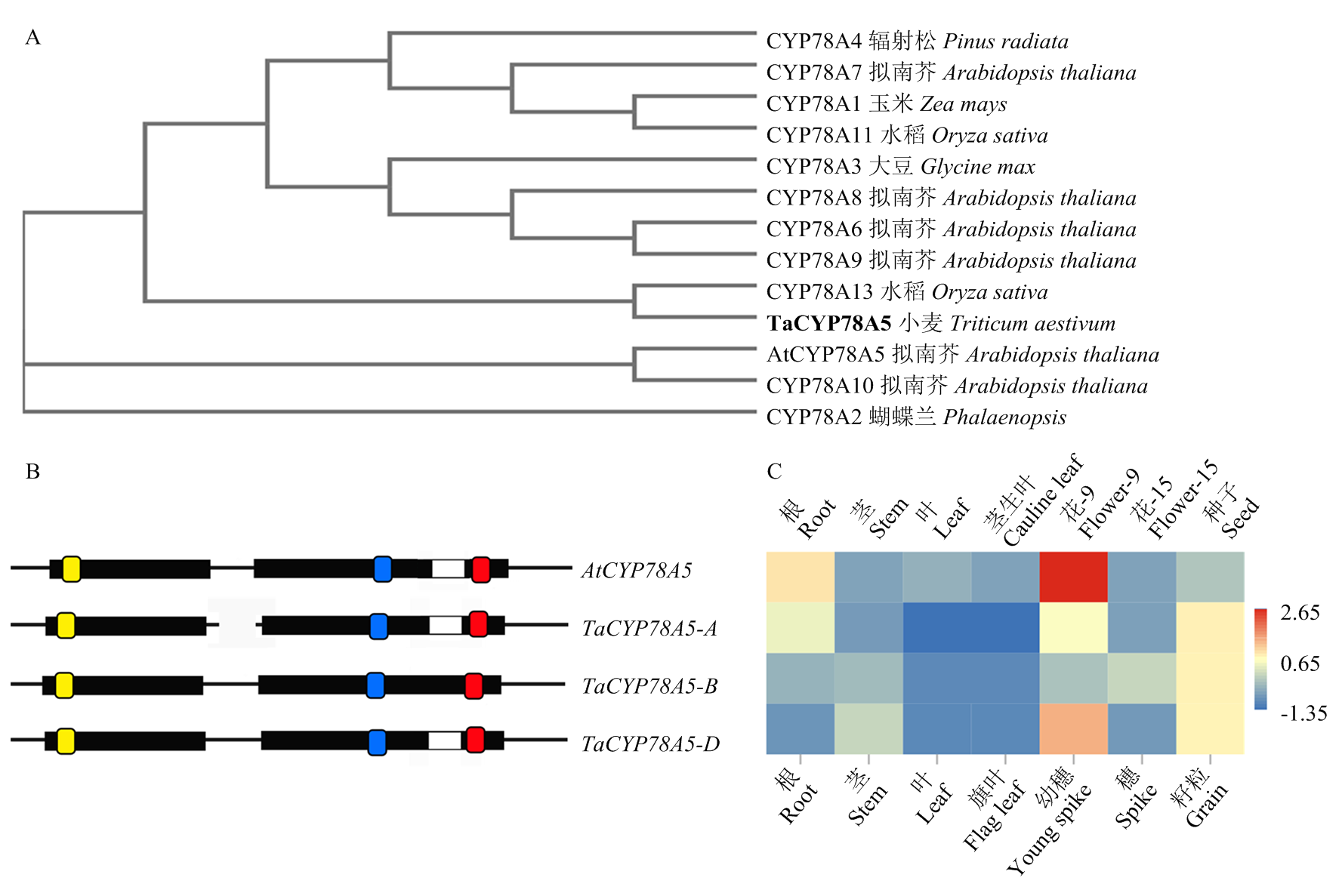
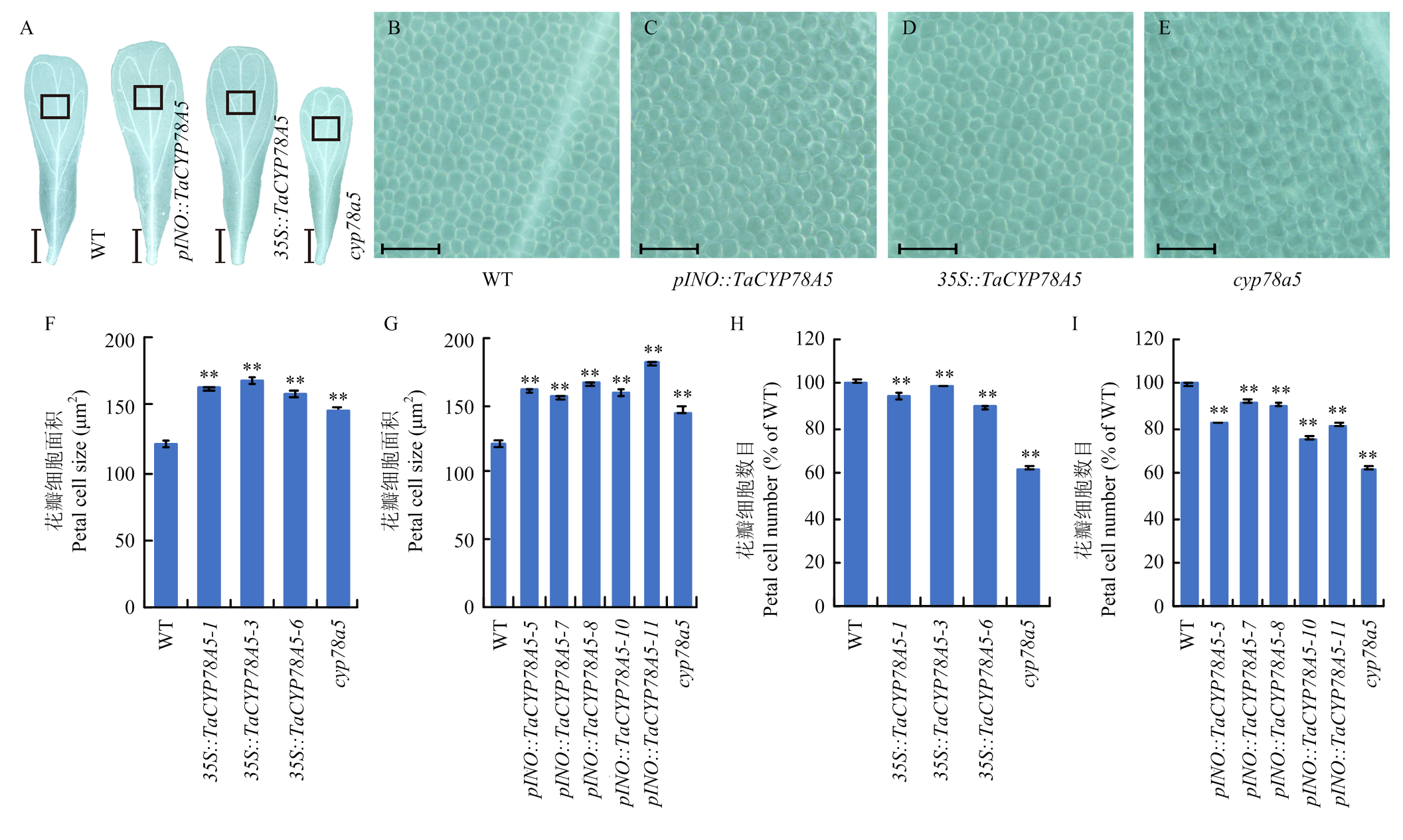
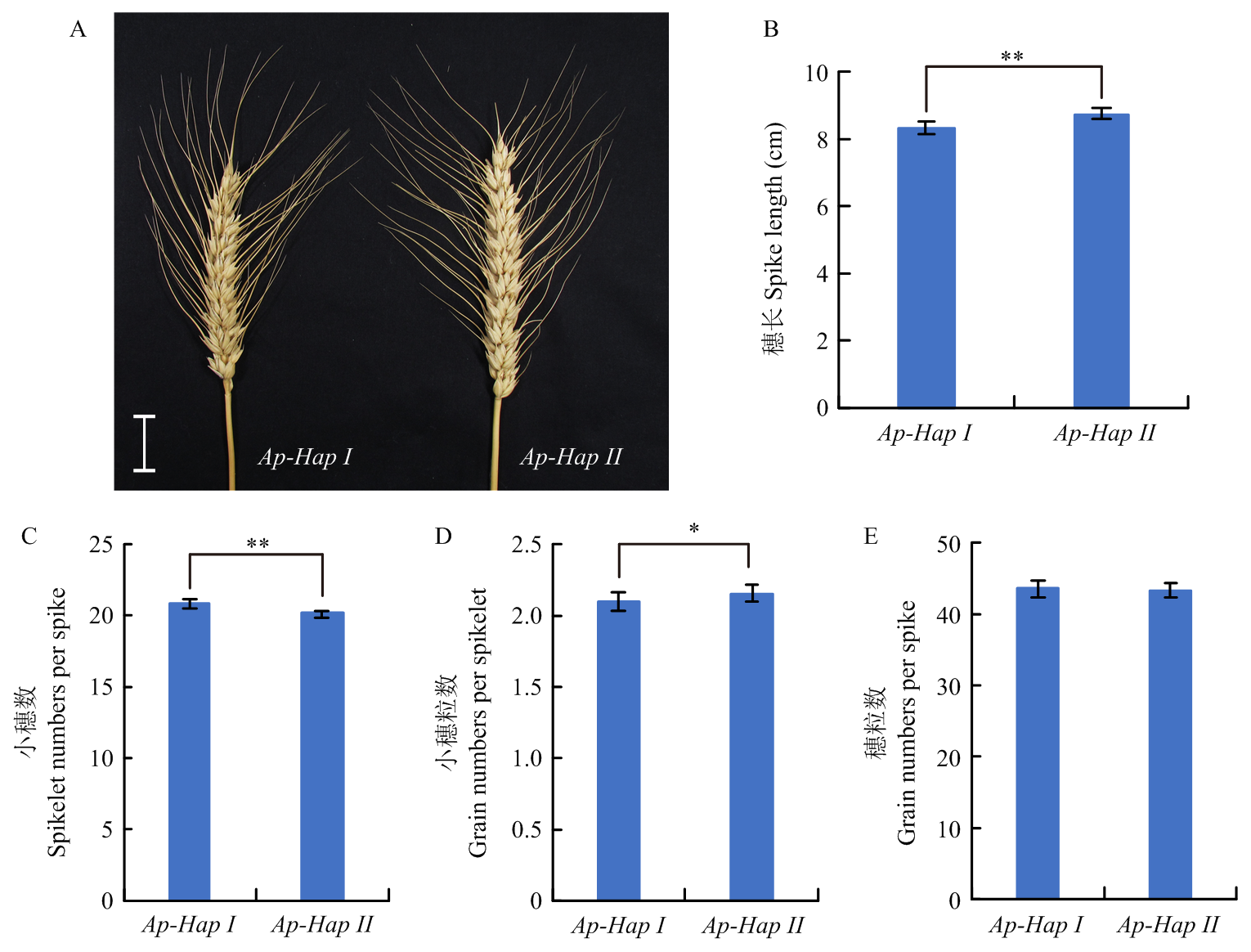
周梦蝶, 李梦瑶, 吴林楠, 郭利建, 王倩, 马猛. 小麦TaKLU基因调控植物株型的研究. 麦类作物学报, 2022, 42(4): 399-406.
|
|
|
|
| [37] |
doi: 10.1046/j.1439-0310.2002.00778.x |
| [38] |
doi: 10.1111/nph.2010.188.issue-2 |
| [39] |
doi: 10.1086/319193 pmid: 18707288 |
| [40] |
doi: 10.2135/cropsci2016.06.0511 |
| [41] |
doi: 10.1093/jxb/erv378 pmid: 26246614 |
| [42] |
doi: 10.1093/jxb/erp019 pmid: 19228817 |
| [43] |
doi: 10.1093/jxb/eru549 pmid: 25609830 |
| [44] |
doi: 10.1146/arplant.1998.49.issue-1 |
| [45] |
|
| [46] |
doi: 10.1101/gad.13.23.3160 |
| [47] |
doi: 10.1086/285413 |
| [48] |
|
| [49] |
doi: 10.1105/tpc.110.081653 |
| [50] |
doi: 10.1007/s00425-012-1639-5 pmid: 22476293 |
| [51] |
doi: 10.1038/nchembio.165 pmid: 19377456 |
| [52] |
|
| [1] | ZHU Qi, JIA ZhenPeng, Tahir SHAH, XU ChenSheng, LI ZhiQi, LÜ HuiShuai, ZHU PengChao, WEI XiaoMin, HUANG DongLin, SUN YanNi, CAO WeiDong, GAO YaJun, WANG ZhaoHui, ZHANG DaBin. Green Manure Crops Combined with Enhanced-Efficiency Products Reduced Greenhouse Gas Emissions and Carbon Footprints in Dryland Wheat Fields [J]. Scientia Agricultura Sinica, 2026, 59(7): 1507-1522. |
| [2] | LI WenHu, LI HaiFeng, DU YuPeng, DING YuLan, LUO YiNuo, LI YuKe, SHE WenTing, ZHANG Feng, TENG Yu, ZHANG SiQi, HUANG Cui, LI XiaoHan, LIU JinShan, WANG ZhaoHui. Regional Differences in Wheat Zinc Uptake and Translocation Responses to Soil Zinc Fertilization [J]. Scientia Agricultura Sinica, 2026, 59(5): 1034-1047. |
| [3] | JIAO WenJuan, HE WanLong, GENG HongWei, BAI Bin, LI JianFeng, CHENG YuKun. Stripe Rust Resistance Evaluation and Molecular Characterization of Yr Genes for 155 Spring Wheat Varieties (Lines) [J]. Scientia Agricultura Sinica, 2026, 59(5): 937-950. |
| [4] | CUI ShiYou, CHEN PengJun, MIAO YuanQing, HAN JiJun, SHEN JunMing. Development and Field Evaluation of Glyphosate-Resistant Wheat Germplasm Generated Through EMS Mutagenesis [J]. Scientia Agricultura Sinica, 2026, 59(4): 723-733. |
| [5] | QIAN Jin, LI YingXue, WU Fang, ZOU XiaoChen. Improved Leaf Phosphorus Content Estimation of Winter Wheat Using Ensemble Hyperspectral Dimensionality Reduction Method [J]. Scientia Agricultura Sinica, 2026, 59(4): 781-792. |
| [6] | KONG Yuan, CUI ShaSha, LI Mei, LI Jian, YANG SiYu, FANG Feng, LIU ShuaiShuai, LIU MingPing, ZENG Yan, GAO XingXiang, BAI LianYang. Spatiotemporal Distribution Dynamics of Five Grass Weed Species Including Lolium multiflorum in Winter Wheat Fields of the Huang- Huai-Hai Region [J]. Scientia Agricultura Sinica, 2026, 59(4): 807-823. |
| [7] | WANG YongSheng, NIU Li, WANG ChangJie, MA LiHua, LIAN XiaoXiao, MENG YaXiong, MA XiaoLe, YAO LiRong, ZHANG Hong, YANG Ke, LI BaoChun, WANG HuaJun, SI ErJing, WANG JunCheng. Genome-Wide Association Study and Candidate Gene Identification for Thousand Grain Weight in Winter Wheat [J]. Scientia Agricultura Sinica, 2026, 59(3): 499-514. |
| [8] | LI XinYi, LI JiaNing, YANG WenPing, XIA Qing, HUO YingRui, HAO ShiHang, HUANG TingMiao, REN YongKang, CHEN Jie, GAO ZhiQiang, YANG ZhenPing. Effects of Post-Anthesis Foliar Zinc Application on Zinc Nutrition in Colored-Grain Wheat [J]. Scientia Agricultura Sinica, 2026, 59(3): 515-527. |
| [9] | XIAN QingLin, XIAO JianKe, GAO AQing, GAO LiChuang, LIU Yang. Effects of Planting Patterns Combined with Soil Moisture Measurement and Supplementary Irrigation on the Yield and Water Use Efficiency of Winter Wheat [J]. Scientia Agricultura Sinica, 2026, 59(3): 589-601. |
| [10] | ZHANG ZhiYong, TAN ShiChao, XIONG ShuPing, MA XinMing, WEI YiHao, WANG XiaoChun. Effects of Annual Water and Nitrogen Optimization on Yield and Nitrogen Migration of Wheat-Maize Rotation System in Irrigation Area of Northern Henan [J]. Scientia Agricultura Sinica, 2026, 59(2): 336-353. |
| [11] | LÜ XuDong, SUN ShiYuan, LI YaNan, LIU YuLong, WANG YanQun, FU Xin, ZHANG JiaYing, NING Peng, PENG ZhengPing. Effects of Intelligent Mechanized Layered Fertilization on Root-Soil Nutrient Distribution and Yield in Wheat Fields [J]. Scientia Agricultura Sinica, 2026, 59(1): 129-146. |
| [12] | LU Hao, ZHANG MingLong, HAN Mei, YAN QingBiao, LI ZhengPeng, YIN Wen, FAN ZhiLong, HU FaLong, CHAI Qiang. Green Manure Returning via Sheep Digest with Nitrogen Fertilizer Reduction are Beneficial to Improve Wheat Yield and Soil Quality at Qinghai-Tibet Plateau [J]. Scientia Agricultura Sinica, 2026, 59(1): 147-160. |
| [13] | YE MeiJin, CHEN JiaTing, ZHOU JieGuang, YIN Li, HU XinRong, LAN YuXin, CHEN Bin, SU LongXing, LIU JiaJun, LIU TianChao, LI XiaoYu, MA Jian. Identification, Validation and Genetic Effect Analysis of Major QTL for Spike Density in Wheat [J]. Scientia Agricultura Sinica, 2026, 59(1): 17-28. |
| [14] | LI YunLi, DIAO DengChao, LIU YaRui, SUN YuChen, MENG XiangYu, WU ChenFang, WANG Yu, WU JianHui, LI ChunLian, ZENG QingDong, HAN DeJun, ZHENG WeiJun. Genome-Wide Association Study of Heat Tolerance at Seedling Stage in A Wheat Natural Population [J]. Scientia Agricultura Sinica, 2025, 58(9): 1663-1683. |
| [15] | PU LiXia, ZHANG JiaRui, YE JianPing, HUANG XiuLan, FAN GaoQiong, YANG HongKun. The Combined Effects of 16, 17-Dihydro Gibberellin A5 and Straw Mulching on Tillering and Grain Yield of Dryland Wheat [J]. Scientia Agricultura Sinica, 2025, 58(9): 1735-1748. |
|
||