Scientia Agricultura Sinica ›› 2024, Vol. 57 ›› Issue (3): 429-441.doi: 10.3864/j.issn.0578-1752.2024.03.001
• CROP GENETICS & BREEDING·GERMPLASM RESOURCES·MOLECULAR GENETICS • Previous Articles Next Articles
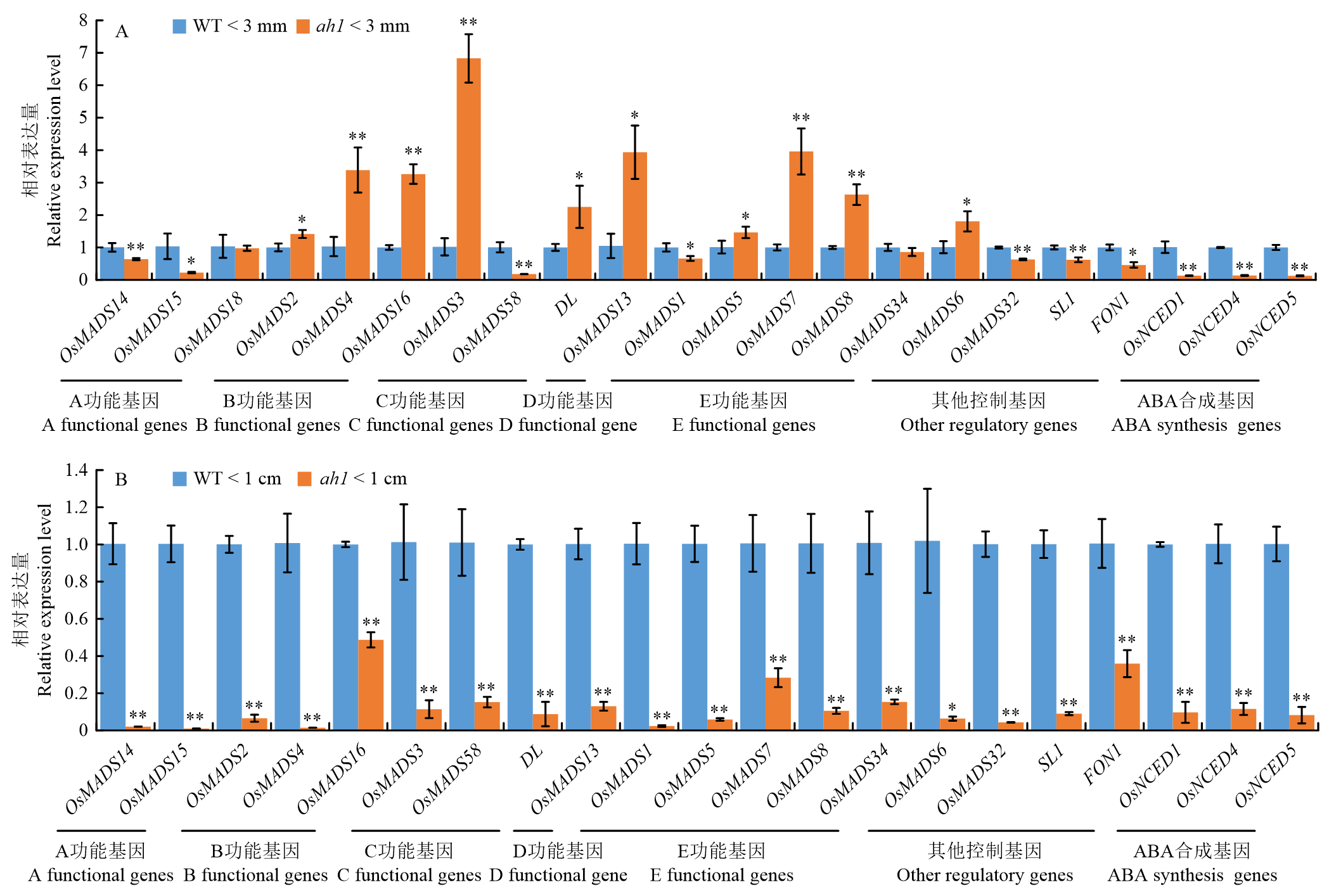
ZHANG BiDong1,2( ), LIN Hong1, ZHU SiYing1, LI ZhongCheng1, ZHUANG Hui1, LI YunFeng1()
), LIN Hong1, ZHU SiYing1, LI ZhongCheng1, ZHUANG Hui1, LI YunFeng1()
| [1] |
doi: 10.1038/353031a0 |
| [2] |
doi: 10.1093/pcp/pcf010 |
| [3] |
pmid: 11832239 |
| [4] |
doi: 10.1093/aob/mcm143 pmid: 17670752 |
| [5] |
pmid: 17616563 |
| [6] |
pmid: 12815719 |
| [7] |
doi: 10.1007/s00497-002-0151-7 |
| [8] |
doi: 10.1371/journal.pgen.1000818 |
| [9] |
doi: 10.1104/pp.104.045039 |
| [10] |
doi: 10.1105/tpc.112.097105 |
| [11] |
doi: 10.1242/dev.00294 |
| [12] |
doi: 10.1023/A:1025401611354 |
| [13] |
doi: 10.1007/s00425-003-1066-8 |
| [14] |
doi: 10.1534/genetics.107.071746 |
| [15] |
doi: 10.1093/pcp/pcn050 |
| [16] |
doi: 10.1093/genetics/165.4.2301 |
| [17] |
doi: 10.1023/a:1006051911291 pmid: 9869408 |
| [18] |
doi: 10.1105/tpc.105.037200 |
| [19] |
doi: 10.1105/tpc.111.087007 pmid: 21810995 |
| [20] |
doi: 10.1093/mp/sst003 |
| [21] |
doi: 10.1105/tpc.018044 |
| [22] |
doi: 10.1111/tpj.2007.52.issue-4 |
| [23] |
|
| [24] |
|
| [25] |
doi: 10.1111/tpj.2005.43.issue-6 |
| [26] |
|
| [27] |
doi: 10.1007/s00425-005-0141-8 |
| [28] |
doi: 10.1007/s11103-005-2161-y |
| [29] |
doi: 10.1016/S1016-8478(23)13260-2 |
| [30] |
doi: 10.1111/j.1365-313X.2009.04101.x pmid: 20003164 |
| [31] |
doi: 10.1104/pp.110.156711 |
| [32] |
doi: 10.1007/s11103-012-9958-2 |
| [33] |
doi: 10.1105/tpc.109.068742 |
| [34] |
doi: 10.1104/pp.112.200980 pmid: 22891238 |
| [35] |
doi: 10.1016/j.ydbio.2005.03.016 |
| [36] |
doi: 10.1111/j.1365-313X.2009.03913.x pmid: 19453444 |
| [37] |
|
| [38] |
doi: 10.1016/j.jplph.2014.06.022 |
| [39] |
pmid: 16511358 |
| [40] |
doi: 10.1007/s00425-021-03802-1 pmid: 34842977 |
| [41] |
doi: 10.1016/j.pestbp.2015.05.006 |
| [42] |
pmid: 16299174 |
| [43] |
doi: 10.1111/j.1365-313X.2011.04871.x pmid: 22126457 |
| [1] | YE MeJin, WU Lei, MD NAHIBUZZAMAN Lohani, YIN Li, HU XinRong, LIU YaXi, JIANG YunFeng, CHEN GuoYue, PU ZhiEn, LI Yang, LI Ting, ZOU YaYa, WU JiaYi, MA Jian. Genome-Wide Association Study-Based Identification of Loci Controlling Mature Embryo Size in Chinese Wheat Landraces and Their Genetic Effects Analysis [J]. Scientia Agricultura Sinica, 2026, 59(6): 1157-1171. |
| [2] | WANG YongSheng, NIU Li, WANG ChangJie, MA LiHua, LIAN XiaoXiao, MENG YaXiong, MA XiaoLe, YAO LiRong, ZHANG Hong, YANG Ke, LI BaoChun, WANG HuaJun, SI ErJing, WANG JunCheng. Genome-Wide Association Study and Candidate Gene Identification for Thousand Grain Weight in Winter Wheat [J]. Scientia Agricultura Sinica, 2026, 59(3): 499-514. |
| [3] | LI YunLi, DIAO DengChao, LIU YaRui, SUN YuChen, MENG XiangYu, WU ChenFang, WANG Yu, WU JianHui, LI ChunLian, ZENG QingDong, HAN DeJun, ZHENG WeiJun. Genome-Wide Association Study of Heat Tolerance at Seedling Stage in A Wheat Natural Population [J]. Scientia Agricultura Sinica, 2025, 58(9): 1663-1683. |
| [4] | ZHUANG LiHua, LUO Lei, ZHAO ChunFang, WANG JiZhong, ZHANG YaDong, HE Lei. Identification and Gene Mapping of Rice Grain Shape Mutant sgd13 [J]. Scientia Agricultura Sinica, 2025, 58(24): 5097-5109. |
| [5] | ZENG YueHui, ZOU WenGuang, ZHAO FuMing, XIAO ChangChun, HUANG JianHong, MA BinLin, YANG WangXing, WEI XinYu, XU XuMing. Map-Based Cloning and Functional Verification of A Novel Split Glume Gene OsSG2 in Rice (Oryza sativa L.) [J]. Scientia Agricultura Sinica, 2025, 58(11): 2062-2080. |
| [6] | ZHANG Ying, SHI TingRui, CAO Rui, PAN WenQiu, SONG WeiNing, WANG Li, NIE XiaoJun. Genome-Wide Association Study of Drought Tolerance at Seedling Stage in ICARDA-Introduced Wheat [J]. Scientia Agricultura Sinica, 2024, 57(9): 1658-1673. |
| [7] | XU Na, TANG Ying, XU ZhengJin, SUN Jian, XU Quan. Genetic Analysis and Candidate Gene Identification on Fertility and Inheritance of Hybrid Sterility of XI and GJ Cross [J]. Scientia Agricultura Sinica, 2024, 57(8): 1417-1429. |
| [8] | HAN XuDong, YANG ChuanQi, ZHANG Qing, LI YaWei, YANG XiaXia, HE JiaTian, XUE JiQuan, ZHANG XingHua, XU ShuTu, LIU JianChao. QTL Mapping and Candidate Gene Screening for Nitrogen Use Efficiency in Maize [J]. Scientia Agricultura Sinica, 2024, 57(21): 4175-4191. |
| [9] | LIU DeLong, LI ShiRu, WANG ChuanXing, GUO ShuQing, MA ZhiXiu, WU YongJiang, HAN HuiBing, LI YuJie, ZHANG PanPan, YANG Pu. Phenotypical Variation and Dynamic QTL Mapping of Plant Height in Foxtail Millet at Different Developmental Stages [J]. Scientia Agricultura Sinica, 2024, 57(18): 3533-3550. |
| [10] | SHANG Hang, CHENG YuKun, REN Yi, GENG HongWei. Genome-Wide Association Analysis of Starch Gelatinization Traits in Winter Wheat [J]. Scientia Agricultura Sinica, 2024, 57(18): 3507-3521. |
| [11] | BAI BingNan, QIAO Dan, GE Qun, LUAN YuJuan, LIU XiaoFang, LU QuanWei, NIU Hao, GONG JuWu, GONG WanKui, ELAMEER ELSAMMAN, YAN HaoLiang, LI JunWen, LIU AiYing, SHI YuZhen, WANG HaiZe, YUAN YouLu. QTN Mining and Candidate Gene Screening of Upland Cotton (Gossypium hirsutum L.) Seed-Related Traits [J]. Scientia Agricultura Sinica, 2024, 57(15): 2901-2913. |
| [12] | YAN LiuHui, ZHONG Qi, MA ZengFeng, WEI MinYi, LIU Chi, QIN YuanYuan, ZHOU XiaoLong, HUANG DaHui, LU YingPing, QIN Gang, ZHANG YueXiong. Identification and Evolutionary Analysis of the Early Heading Gene OsEHD8 in Common Wild Rice (Oryza rufipogon Giff.) [J]. Scientia Agricultura Sinica, 2024, 57(14): 2703-2716. |
| [13] | ZHANG YuMei, DING WenTao, LAN XinLong, LI QingHua, HU RunFang, GUO Na, LIN GuoQiang, ZHAO JinMing. Genome Wide Association Analysis of Soluble Sugar Content in Fresh Seeds of Soybean Landraces [J]. Scientia Agricultura Sinica, 2024, 57(11): 2079-2091. |
| [14] | LI ShengYou, WANG ChangLing, YAN ChunJuan, ZHANG LiJun, SUN XuGang, CAO YongQiang, WANG WenBin, SONG ShuHong. Evaluation of Drought Resistance in Soybean Germplasm and Identification of Candidate Drought-Resistant Genes [J]. Scientia Agricultura Sinica, 2024, 57(10): 1857-1869. |
| [15] | ZHU HongHui, LI YingZi, GAO YuanZhuo, LIN Hong, WANG ChengYang, YAN ZiYi, PENG HanPing, LI TianYe, XIONG Mao, LI YunFeng. Map-Based Cloning of the SHORT AND WIDEN GRAIN 1 Gene in Rice (Oryza sativa L.) [J]. Scientia Agricultura Sinica, 2023, 56(7): 1260-1274. |
|
||