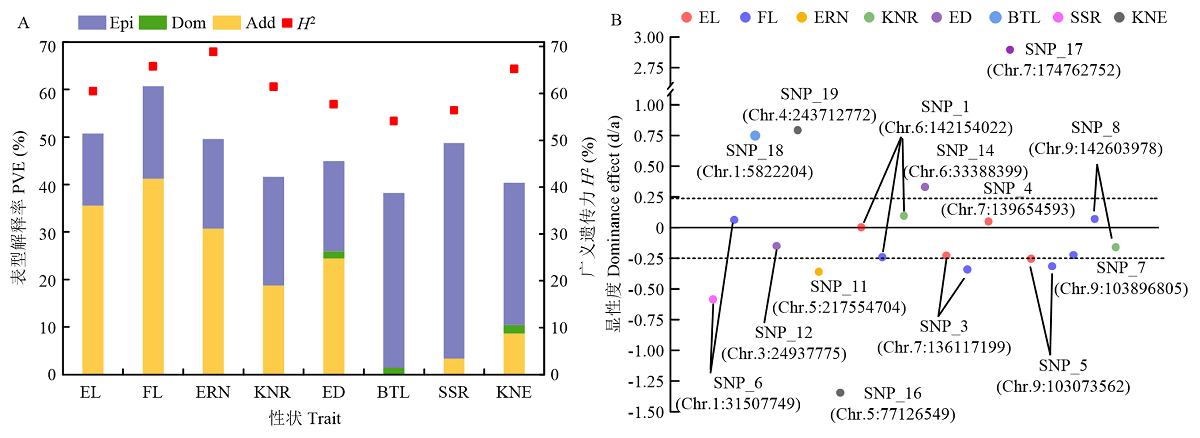
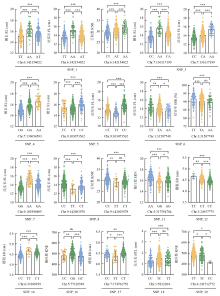
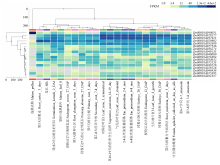
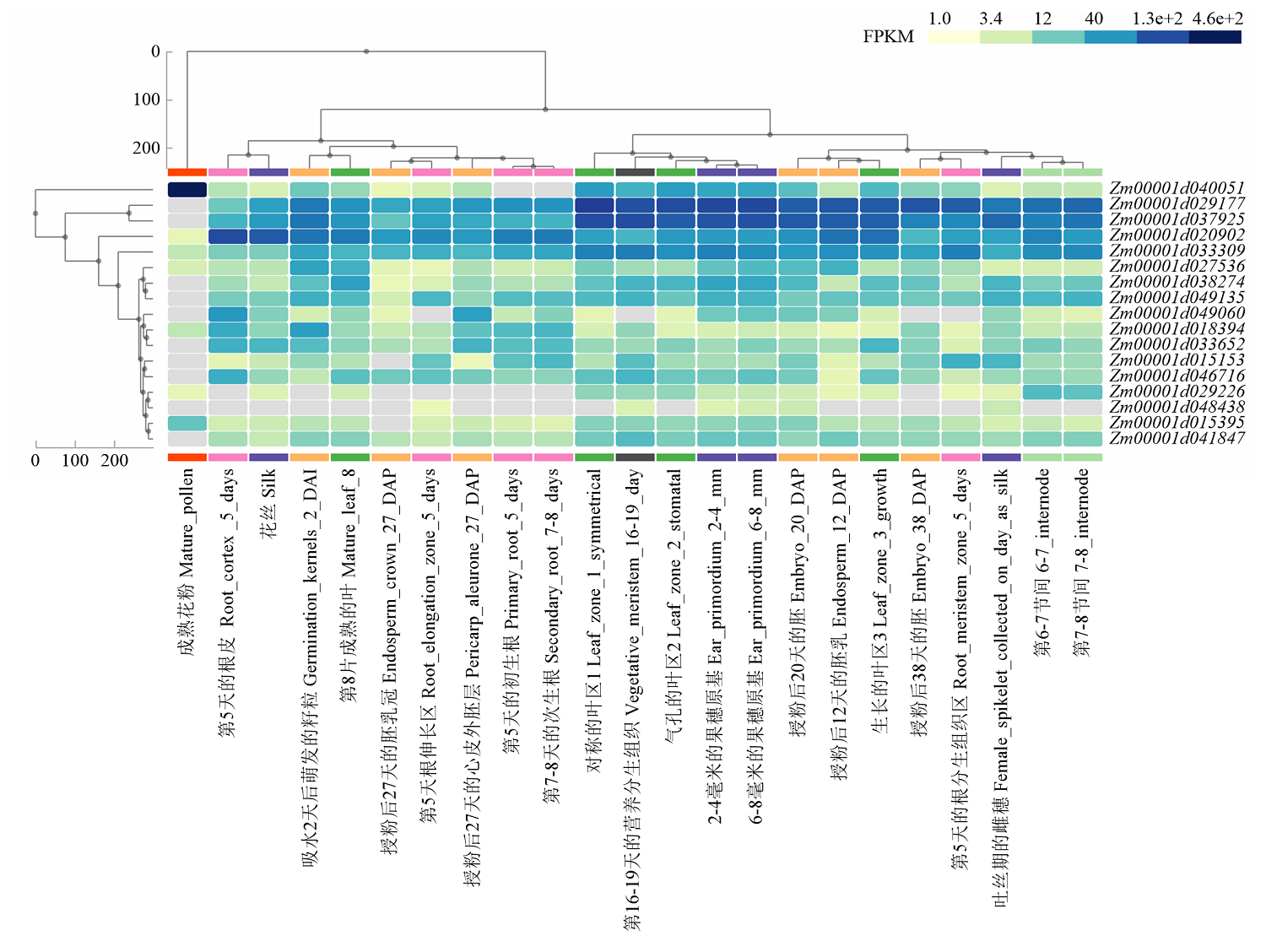
Scientia Agricultura Sinica ›› 2022, Vol. 55 ›› Issue (13): 2485-2499.doi: 10.3864/j.issn.0578-1752.2022.13.001
• CROP GENETICS & BREEDING·GERMPLASM RESOURCES·MOLECULAR GENETICS • Previous Articles Next Articles
LI Ting( ),DONG Yuan,ZHANG Jun,FENG ZhiQian,WANG YaPeng,HAO YinChuan,ZHANG XingHua,XUE JiQuan(),XU ShuTu()
),DONG Yuan,ZHANG Jun,FENG ZhiQian,WANG YaPeng,HAO YinChuan,ZHANG XingHua,XUE JiQuan(),XU ShuTu()
| [1] | 赵久然, 郭景伦, 郭强, 尉德明, 肖必祥, 卢柏山. 玉米不同品种基因型穗粒数及其构成因素相关分析的研究. 北京农业科学, 1997, 15(6): 1-2. |
| ZHAO J R, GUO J L, GUO Q, WEI D M, XIAO B X, LU B S. Correlation analysis of grain number per ear and its components in different genotypes of maize. Beijing Agriculture Sciences, 1997, 15(6): 1-2. (in Chinese) | |
| [2] | 何坤辉, 常立国, 李亚楠, 渠建洲, 崔婷婷, 徐淑兔, 薛吉全, 刘建超. 供氮和不供氮条件下玉米穗部性状的QTL定位. 植物营养与肥料学报, 2017, 23(1): 91-100. |
| HE K H, CHANG L G, LI Y N, QU J Z, CUI T T, XU S T, XUE J Q, LIU J C. QTL mapping of ear traits of maize with and without N input. Journal of Plant Nutrition and Fertilizer, 2017, 23(1): 91-100. (in Chinese) | |
| [3] |
张焕欣, 翁建峰, 张晓聪, 刘昌林, 雍洪军, 郝转芳, 李新海. 玉米穗行数全基因组关联分析. 作物学报, 2014, 40(1): 1-6.
doi: 10.3724/SP.J.1006.2014.00001 |
|
ZHANG H X, WENG J F, ZHANG X C, LIU C L, YONG H J, HAO Z F, LI X H. Genome-wide association analysis of kernel row number in maize. Acta Agronomica Sinica, 2014, 40(1): 1-6. (in Chinese)
doi: 10.3724/SP.J.1006.2014.00001 |
|
| [4] |
吴律, 代力强, 董青松, 施婷婷, 王丕武. 玉米行粒数的全基因组关联分析. 作物学报, 2017, 43(10): 1559-1564.
doi: 10.3724/SP.J.1006.2017.01559 |
|
WU L, DAI L Q, DONG Q S, SHI T T, WANG P W. Genome-wide association analysis of kernel number per row in maize. Acta Agronomica Sinica, 2017, 43(10): 1559-1564. (in Chinese)
doi: 10.3724/SP.J.1006.2017.01559 |
|
| [5] |
马娟, 曹言勇, 李会勇. 玉米穗轴粗全基因组关联分析. 作物学报, 2021, 47(7): 1228-1238.
doi: 10.3724/SP.J.1006.2021.03048 |
|
MA J, CAO Y Y, LI H Y. Genome-wide association study of ear cob diameter in maize. Acta Agronomica Sinica, 2021, 47(7): 1228-1238. (in Chinese)
doi: 10.3724/SP.J.1006.2021.03048 |
|
| [6] |
YANG L, LI T, TIAN X K, YANG B B, LAO Y H, WANG Y H, ZHANG X H, XUE J Q, XU S T. Genome-wide association study (GWAS) reveals genetic basis of ear-related traits in maize. Euphytica, 2020, 216(11): 1-13.
doi: 10.1007/s10681-019-2539-6 |
| [7] | 钱佳翼, 柳俊, 方圆, 杜勇成, 杨天天, 李鹏程, 杨泽峰, 徐辰武. 玉米穗型和粒型性状的GWAS及其关联位点驯化和改良分析. 玉米科学, 2020, 28(6): 45-51. |
| QIAN J Y, LIU J, FANG Y, DU Y C, YANG T T, LI P C, YANG Z F, XU C W. Genome-wide association study on ear-and kernel-related traits in maize and the analysis of domestication and improvement of the associated loci. Journal of Maize Sciences, 2020, 28(6): 45-51. (in Chinese) | |
| [8] |
XIAO Y J, TONG H, YANG X H, XU S Z, PAN Q C, QIAO F, RAIHAN M S, LUO Y, LIU H J, ZHANG X H, YANG N, WANG X Q, DENG M, JIN M L, ZHAO L J, LUO X, ZHOU Y, LI X, LIU J, ZHAN W, LIU N N, WANG H, CHEN G S, CAI Y, XU G, WANG W D, ZHENG D B, YAN J B. Genome-wide dissection of the maize ear genetic architecture using multiple populations. New Phytologist, 2016, 210(3): 1095-1110.
doi: 10.1111/nph.13814 |
| [9] | 殷芳冰, 王成, 龙艳, 董振营, 万向元. 玉米雌穗性状遗传分析与形成机制. 中国生物工程杂志, 2022, 41(12): 30-46. |
| YIN F B, WANG C, LONG Y, DONG Z Y, WAN X Y. Genetic architecture and formation mechanism of female ear traits in maize. China Biotechnology, 2022, 41(12): 30-46. (in Chinese) | |
| [10] |
LI M F, ZHONG W S, YANG F, ZHANG Z X. Genetic and molecular mechanisms of quantitative trait loci controlling maize inflorescence architecture. Plant and Cell Physiology, 2018, 59(3): 448-457.
doi: 10.1093/pcp/pcy022 |
| [11] |
CHUCK G S, BROWN P J, MEELEY R, HAKE S. Maize SBP-box transcription factors unbranched2 and unbranched3 affect yield traits by regulating the rate of lateral primordia initiation. Proceedings of the National Academy of Sciences of the USA, 2014, 111(52): 18775-18780.
doi: 10.1073/pnas.1407401112 |
| [12] |
JIA H T, LI M F, LI W Y, LIU L, JIAN Y N, YANG Z X, SHEN X M, NING Q, DU Y F, ZHAO R, JACKSON D, YANG X H, ZHANG Z X. A serine/threonine protein kinase encoding gene KERNEL NUMBER PER ROW6 regulates maize grain yield. Nature Communications, 2020, 11(1): 1-11.
doi: 10.1038/s41467-019-13993-7 |
| [13] |
NING Q, JIAN Y N, DU Y F, LI Y F, SHEN X M, JIA H T, ZHAO R, ZHAN J M, YANG F, JACKSON D, LIU L, ZHANG Z X. An ethylene biosynthesis enzyme controls quantitative variation in maize ear length and kernel yield. Nature Communications, 2021, 12(1): 1-10.
doi: 10.1038/s41467-020-20314-w |
| [14] |
WANG J, LIN Z L, ZHANG X, LIU H Q, ZHOU L N, ZHONG S Y, LI Y, ZHU C, LIN Z W. krn1, a major quantitative trait locus for kernel row number in maize. New Phytologist, 2019, 223(3): 1634-1646.
doi: 10.1111/nph.15890 |
| [15] |
BOMMERT P, NAGASAWA N S, JACKSON D. Quantitative variation in maize kernel row number is controlled by the FASCIATED EAR2 locus. Nature Genetics, 2013, 45(3): 334-337.
doi: 10.1038/ng.2534 |
| [16] |
KNAPP S J, STROUP W W, ROSS W M. Exact confidence intervals for heritability on a progeny mean basis 1. Crop Science, 1985, 25(1): 192-194.
doi: 10.2135/cropsci1985.0011183X002500010046x |
| [17] |
MURRAY M G, THOMPSON W F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Research, 1980, 8(19): 4321-4326.
doi: 10.1093/nar/8.19.4321 |
| [18] |
LI T, QU J Z, WANG Y H, CHANG L G, HE K H, GUO D W, ZHANG X H, XU S T, XUE J Q. Genetic characterization of inbred lines from Shaan A and B groups for identifying loci associated with maize grain yield. BMC Genetics, 2018, 19(1): 1-12.
doi: 10.1186/s12863-017-0594-3 |
| [19] |
PURCELL S, NEALE B, TODD-BROWN K, THOMAS L, FERREIRA M A, BENDER D, MALLER J, SKLAR P, De BAKKER P I, DALY M J, SHAM P C. PLINK: A tool set for whole-genome association and population-based linkage analyses. The American Journal of Human Genetics, 2007, 81(3): 559-575.
doi: 10.1086/519795 |
| [20] |
BRADBURY P J, ZHANG Z, KROON D E, CASSTEVENS T M, RAMDOSS Y, BUCKLER E S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics, 2007, 23(19): 2633-2635.
doi: 10.1093/bioinformatics/btm308 |
| [21] |
JIANG Y, SCHMIDT R H, ZHAO Y S, REIF J C. A quantitative genetic framework highlights the role of epistatic effects for grain-yield heterosis in bread wheat. Nature Genetics, 2017, 49(12): 1741-1746.
doi: 10.1038/ng.3974 |
| [22] |
GAO X Y, STARMER J, MARTIN E R. A multiple testing correction method for genetic association studies using correlated single nucleotide polymorphisms. Genetic Epidemiology, 2008, 32(4): 361-369.
doi: 10.1002/gepi.20310 |
| [23] | HUANG X H, YANG S H, GONG J Y, ZHAO Y, FENG Q, GONG H, LI W J, ZHAN Q L, CHENG B Y, XIA J H, CHEN N, HAO Z N, LIU K Y, ZHU C R, HUANG T, ZHAO Q, ZHANG L, FAN D L, ZHOU C C, LU Y Q, WENG Q J, WANG Z X, LI J Y, HAN B. Genomic analysis of hybrid rice varieties reveals numerous superior alleles that contribute to heterosis. Nature Communications, 2015, 6(1): 1-9. |
| [24] | WANG K, LI M, HAKONARSON H. ANNOVAR: Functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Research, 2010, 38(16): e164-e164. |
| [25] | 黄远樟, 刘来福. 作物数量遗传学基础——六、配合力: 不完全双列杂交. 遗传, 1980(2): 43-46. |
| HUANG Y Z, LIU L F. The basis of quantitative genetics in crops: Ⅵ. Combining ability: Incomplete diallel cross. Hereditas, 1980(2): 43-46. (in Chinese) | |
| [26] | 刘文童, 监立强, 郭晋杰, 赵永锋, 黄亚群, 陈景堂, 祝丽英. 玉米穗部性状及其一般配合力的关联分析. 植物遗传资源学报, 2020, 21(3): 706-715. |
| LIU W T, JIAN L Q, GUO J J, ZHAO Y F, HUANG Y Q, CHEN J T, ZHU L Y. Association analysis of ear-related traits and their general combining ability in maize. Journal of Plant Genetic Resources, 2020, 21(3): 706-715. (in Chinese) | |
| [27] |
CHEN J X, ZHOU H, XIE W B, XIA D, GAO G J, ZHANG Q L, WANG G W, LIAN X M, XIAO J H, HE Y Q. Genome-wide association analyses reveal the genetic basis of combining ability in rice. Plant Biotechnology Journal, 2019, 17(11): 2211-2222.
doi: 10.1111/pbi.13134 |
| [28] |
LI G L, DONG Y, ZHAO Y S, TIAN X K, WÜRSCHUM T, XUE J Q, CHEN S J, REIF J C, XU S T, LIU W X. Genome-wide prediction in a hybrid maize population adapted to Northwest China. The Crop Journal, 2020, 8(5): 830-842.
doi: 10.1016/j.cj.2020.04.006 |
| [29] | SHULL G H. The composition of a field of maize. Journal of Heredity, 1908(1): 296-301. |
| [30] | 王晖. 玉米全基因组关联分析多杂种群体的构建及其杂种优势和配合力的遗传分析[D]. 北京: 中国农业科学院, 2017. |
| WANG H. Development of a maize multiple-hybrid population for genome-wide association studies and genetic analysis of heterosis and combining ability[D]. Beijing: Chinese Academy of Agricultural Sciences, 2017. (in Chinese) | |
| [31] |
LIU H J, WANG Q, CHEN M J, DING Y H, YANG X R, LIU J, LI X H, ZHOU C C, TIAN Q L, LU Y Q, FAN D L, SHI J P, ZHANG L, KANG C B, SUN M F, LI F Y, WU Y J, ZHANG Y Z, LIU B S, ZHAO X Y, FENG Q, YANG J L, HAN B, LAI J S, ZHANG X S, HUANG X H. Genome-wide identification and analysis of heterotic loci in three maize hybrids. Plant Biotechnology Journal, 2020, 18(1): 185-194.
doi: 10.1111/pbi.13186 |
| [32] |
LU M, XIE C X, LI X H, HAO Z F, LI M S, WENG J F, ZHANG D G, BAI L, ZHANG S H. Mapping of quantitative trait loci for kernel row number in maize across seven environments. Molecular Breeding, 2011, 28(2): 143-152.
doi: 10.1007/s11032-010-9468-3 |
| [33] |
LIU L, DU Y F, HUO D G, WANG M, SHEN X M, YUE B, QIU F Z, ZHENG Y L, YAN J B, ZHANG Z X. Genetic architecture of maize kernel row number and whole genome prediction. Theoretical and Applied Genetics, 2015, 128(11): 2243-2254.
doi: 10.1007/s00122-015-2581-2 |
| [34] |
XIAO Y J, JIANG S Q, CHENG Q, WANG X Q, YAN J, ZHANG R Y, QIAO F, MA C, LUO J Y, LI W Q, LIU H J, YANG W Y, SONG W H, MENG Y J, WARBURTON M L, ZHAO J R, WANG X F, YAN J B. The genetic mechanism of heterosis utilization in maize improvement. Genome Biology, 2021, 22(1): 1-29.
doi: 10.1186/s13059-020-02207-9 |
| [35] |
YAN J B, TANG H, HUANG Y Q, ZHENG Y L, LI J S. Quantitative trait loci mapping and epistatic analysis for grain yield and yield components using molecular markers with an elite maize hybrid. Euphytica, 2006, 149(1): 121-131.
doi: 10.1007/s10681-005-9060-9 |
| [36] | MA J, CAO Y Y. Genetic dissection of grain yield of maize and yield-related traits through association mapping and genomic prediction. Frontiers in Plant Science, 2021, 12: 1377. |
| [37] |
XUE S, BRADBURY P J, CASSTEVENS T, HOLLAND J B. Genetic architecture of domestication-related traits in maize. Genetics, 2016, 204(1): 99-113.
doi: 10.1534/genetics.116.191106 |
| [38] |
LI T, QU J Z, TIAN X K, LAO Y H, WEI N N, WANG Y H, HAO Y C, ZHANG X H, XUE J Q, XU S T. Identification of ear morphology genes in maize (Zea mays L.) using selective sweeps and association mapping. Frontiers in Genetics, 2020, 11: 747.
doi: 10.3389/fgene.2020.00747 |
| [39] |
XU C, ZHANG H, SUN J, GUO Z, ZOU C, LI W, XIE C, HUANG C, XU R, LIAO H, WANG J X, XU X J, WANG S H, XU Y B. Genome-wide association study dissects yield components associated with low-phosphorus stress tolerance in maize. Theoretical and Applied Genetics, 2018, 131(8): 1699-1714.
doi: 10.1007/s00122-018-3108-4 |
| [40] |
WANG Z J, YUAN T T, YUAN C, NIU Y Q, SUN D Y, CUI S J. LFR, which encodes a novel nuclear-localized Armadillo-repeat protein, affects multiple developmental processes in the aerial organs in Arabidopsis. Plant Molecular Biology, 2009, 69(1): 121-131.
doi: 10.1007/s11103-008-9411-8 |
| [41] |
YU X M, JIANG L L, WU R, MENG X C, ZHANG A, LI N, XIA Q, QI X, PANG J S, XU Z Y, LIU B. The core subunit of a chromatin-remodeling complex, ZmCHB101, plays essential roles in maize growth and development. Scientific Reports, 2016, 6(1): 1-13.
doi: 10.1038/s41598-016-0001-8 |
| [42] |
PAUTLER M, EVELAND A L, LARUE T, YANG F, WEEKS R, LUNDE C, JE B I, MEELEY R, KOMATSU M, VOLLBRECHT E, SAKAI H, JACKSON D. FASCIATED EAR4 encodes a bZIP transcription factor that regulates shoot meristem size in maize. The Plant Cell, 2015, 27(1): 104-120.
doi: 10.1105/tpc.114.132506 |
| [43] |
NI S, LI Z Z, YING J, ZHANG J C, CHEN H Q. Decreased Spikelets 4 encoding a novel tetratricopeptide repeat domain-containing protein is involved in DNA repair and spikelet number determination in rice. Genes, 2019, 10(3): 214.
doi: 10.3390/genes10030214 |
| [44] |
GAO H R, GORDON-KAMM W J, LYZNIK L A. ASF/SF2-like maize pre-mRNA splicing factors affect splice site utilization and their transcripts are alternatively spliced. Gene, 2004, 339: 25-37.
doi: 10.1016/j.gene.2004.06.047 |
| [45] |
van BUEREN E L, BACKES G, De VRIEND H, ØSTERGÅRD H. The role of molecular markers and marker assisted selection in breeding for organic agriculture. Euphytica, 2010, 175(1): 51-64.
doi: 10.1007/s10681-010-0169-0 |
| [1] | ZHANG Ying, SHI TingRui, CAO Rui, PAN WenQiu, SONG WeiNing, WANG Li, NIE XiaoJun. Genome-Wide Association Study of Drought Tolerance at Seedling Stage in ICARDA-Introduced Wheat [J]. Scientia Agricultura Sinica, 2024, 57(9): 1658-1673. |
| [2] | FAN Hong, YIN Wen, HU FaLong, FAN ZhiLong, ZHAO Cai, YU AiZhong, HE Wei, SUN YaLi, WANG Feng, CHAI Qiang. Compensation Potential of Dense Planting on Nitrogen Reduction in Maize Yield in Oasis Irrigation Area [J]. Scientia Agricultura Sinica, 2024, 57(9): 1709-1721. |
| [3] | XU Na, TANG Ying, XU ZhengJin, SUN Jian, XU Quan. Genetic Analysis and Candidate Gene Identification on Fertility and Inheritance of Hybrid Sterility of XI and GJ Cross [J]. Scientia Agricultura Sinica, 2024, 57(8): 1417-1429. |
| [4] | WANG ChengZe, ZHANG Yan, FU Wei, JIA JingZhe, DONG JinGao, SHEN Shen, HAO ZhiMin. Bioinformatics and Expression Pattern Analysis of Maize ACO Gene Family [J]. Scientia Agricultura Sinica, 2024, 57(7): 1308-1318. |
| [5] | GAO ChenXi, HAO LuYang, HU Yue, LI YongXiang, ZHANG DengFeng, LI ChunHui, SONG YanChun, SHI YunSu, WANG TianYu, LI Yu, LIU XuYang. Analysis of Transposable Element Associated Epigenetic Regulation under Drought in Maize [J]. Scientia Agricultura Sinica, 2024, 57(6): 1034-1048. |
| [6] | ZHAO KaiNan, DING Hao, LIU AKang, JIANG ZongHao, CHEN GuangZhou, FENG Bo, WANG ZongShuai, LI HuaWei, SI JiSheng, ZHANG Bin, BI XiangJun, LI Yong, LI ShengDong, WANG FaHong. Nitrogen Fertilizer Reduction and Postponing for Improving Plant Photosynthetic Physiological Characteristics to Increase Wheat- Maize and Annual Yield and Economic Return [J]. Scientia Agricultura Sinica, 2024, 57(5): 868-884. |
| [7] | WANG Yu, ZHANG YuPeng, ZHU GuanYa, LIAO HangXi, HOU WenFeng, GAO Qiang, WANG Yin. Effects of Localized Nitrogen Supply on Plant Growth and Water and Nitrogen Use Efficiencies of Maize Seedling Under Drought Stress [J]. Scientia Agricultura Sinica, 2024, 57(5): 919-934. |
| [8] | GAO ShangJie, LIU XingRen, LI YingChun, LIU XiaoWan. Effects of Biochar and Straw Return on Greenhouse Gas Emissions and Global Warming Potential in the Farmland [J]. Scientia Agricultura Sinica, 2024, 57(5): 935-949. |
| [9] | LI QianChuan, XU ShiWei, ZHANG YongEn, ZHUANG JiaYu, LI DengHua, LIU BaoHua, ZHU ZhiXun, LIU Hao. Stacking Ensemble Learning Modeling and Forecasting of Maize Yield Based on Meteorological Factors [J]. Scientia Agricultura Sinica, 2024, 57(4): 679-697. |
| [10] | ZHANG BiDong, LIN Hong, ZHU SiYing, LI ZhongCheng, ZHUANG Hui, LI YunFeng. Identification and Candidate Gene Analysis of the ABNORMAL HULL 1 (ah1) Mutant in Rice (Oryza sativa L.) [J]. Scientia Agricultura Sinica, 2024, 57(3): 429-441. |
| [11] | SHI DeYang, LI YanHong, WANG FeiFei, XIA DeJun, JIAO YanLin, SUN NiNa, ZHAO Jian. Regulation Effects of Line-Spacing Expansion and Row-Spacing Shrinkage on Dry Matter and Nutrient Accumulation and Transport of Summer Maize Under High Plant Density [J]. Scientia Agricultura Sinica, 2024, 57(23): 4658-4672. |
| [12] | CAO WenZhuo, YU ZhenWen, ZHANG YongLi, ZHANG Zhen, SHI Yu, WANG YongJun. The Difference of Grain Starch Accumulation Dynamics and Yield Formation of Spring Maize Under Different Nitrogen Application Rates in Black Soil [J]. Scientia Agricultura Sinica, 2024, 57(22): 4431-4443. |
| [13] | DONG KuiJun, ZHANG YiTao, LIU HanWen, ZHANG JiZong, WANG WeiJun, WEN YanChen, LEI QiuLiang, WEN HongDa. Effects of Nitrogen Reduction Application of Summer Maize- Soybean Intercropping on Agronomic Traits and Economic Benefits as well as Its Yield of Subsequent Wheat [J]. Scientia Agricultura Sinica, 2024, 57(22): 4495-4506. |
| [14] | HAN XuDong, YANG ChuanQi, ZHANG Qing, LI YaWei, YANG XiaXia, HE JiaTian, XUE JiQuan, ZHANG XingHua, XU ShuTu, LIU JianChao. QTL Mapping and Candidate Gene Screening for Nitrogen Use Efficiency in Maize [J]. Scientia Agricultura Sinica, 2024, 57(21): 4175-4191. |
| [15] | GUO Ya, REN Hao, WANG HongZhang, ZHANG JiWang, ZHAO Bin, REN BaiZhao, LIU Peng. High Temperature and Drought Combined Stress Inhibited Photosystem Ⅱ Performance and Decreased Grain Yield of Summer Maize [J]. Scientia Agricultura Sinica, 2024, 57(21): 4205-4220. |
|
||