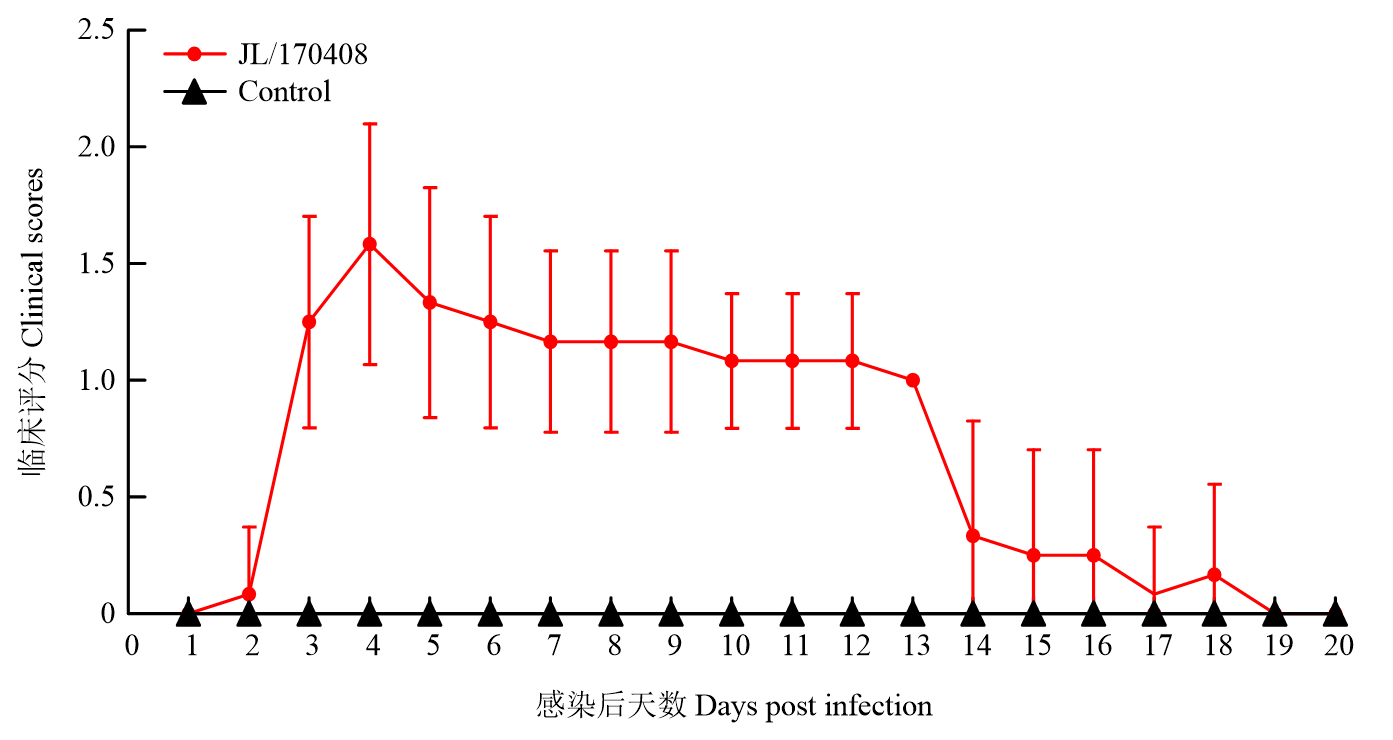
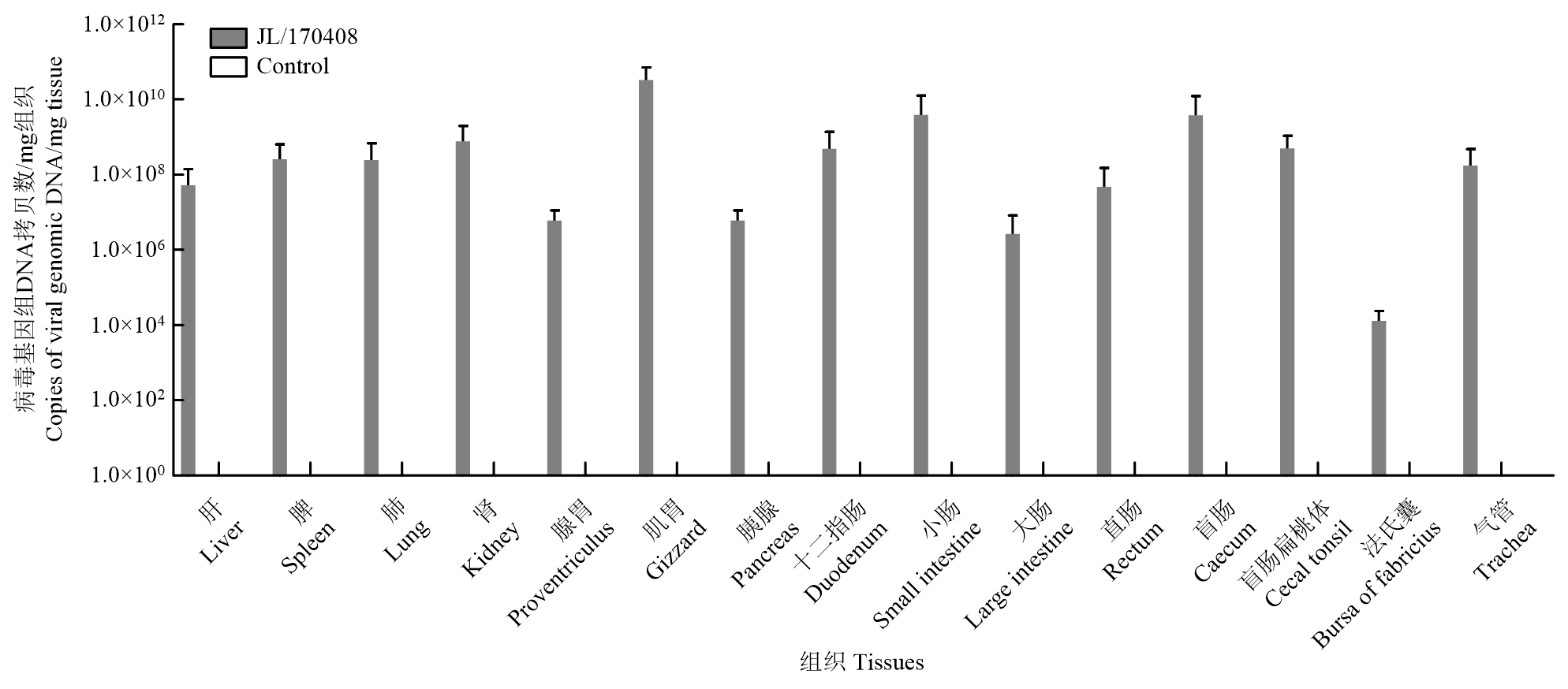
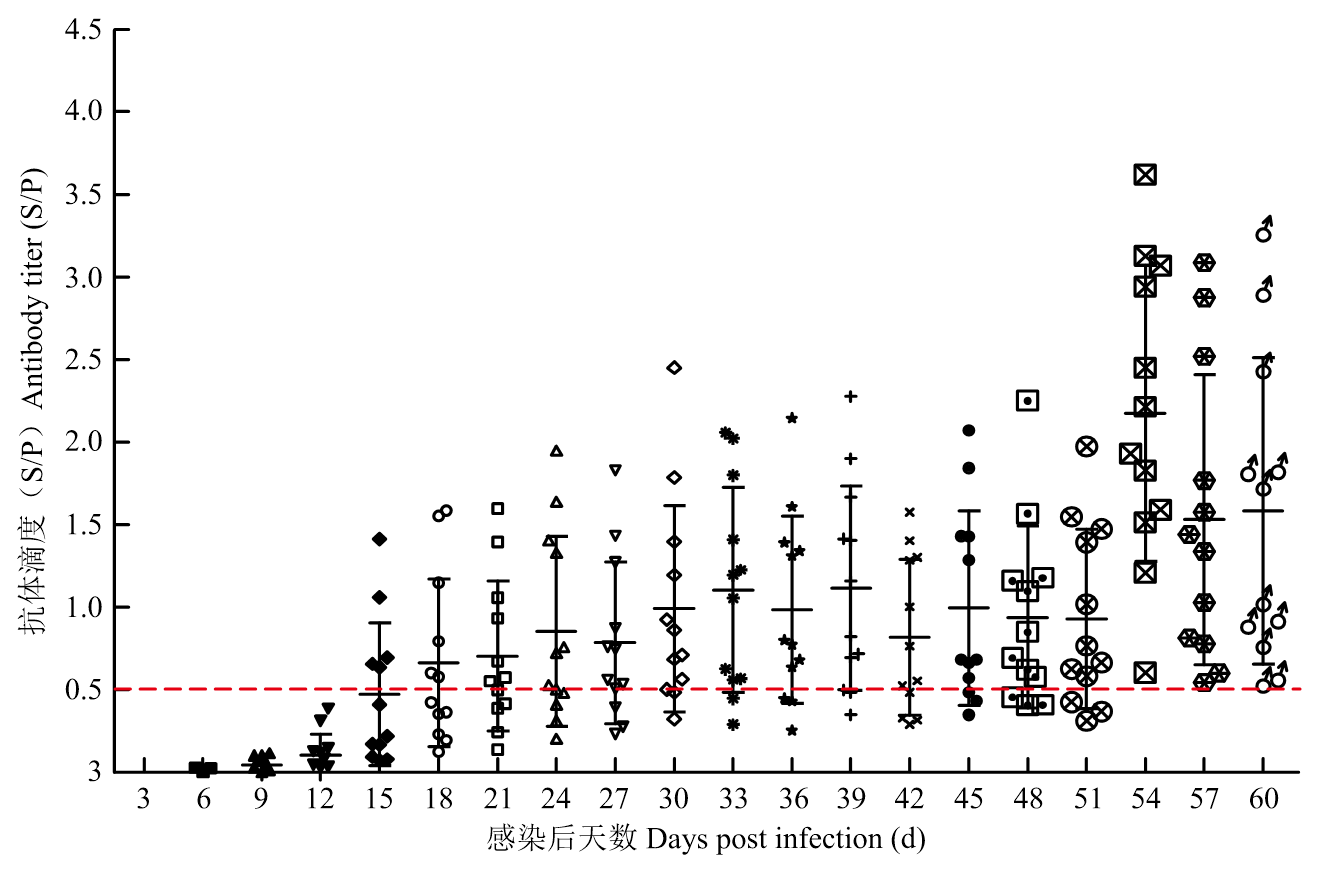
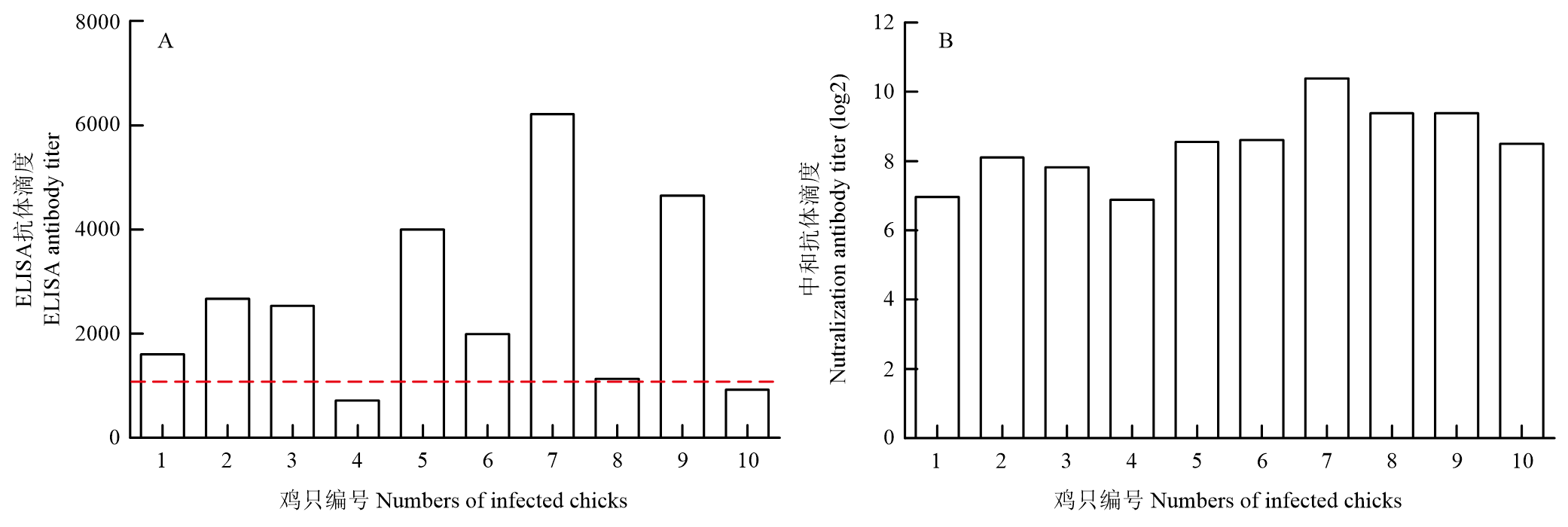
Scientia Agricultura Sinica ›› 2023, Vol. 56 ›› Issue (16): 3226-3236.doi: 10.3864/j.issn.0578-1752.2023.16.014
• ANIMAL SCIENCE·VETERINARY SCIENCE • Previous Articles Next Articles
LI HuiXin( ), SONG WenPing, HAN ZongXi, LIU ShengWang()
), SONG WenPing, HAN ZongXi, LIU ShengWang()
| [1] |
|
| [2] |
doi: 10.1080/03079450050045440 pmid: 19184805 |
| [3] |
doi: 10.1099/vir.0.030064-0 |
| [4] |
doi: 10.1016/j.vetmic.2011.11.003 |
| [5] |
|
| [6] |
pmid: 5896495 |
| [7] |
doi: 10.1637/7577-040106R.1 pmid: 17274293 |
| [8] |
pmid: 22545531 |
| [9] |
doi: 10.1007/s13337-013-0183-7 pmid: 24426318 |
| [10] |
doi: 10.2307/1588976 |
| [11] |
pmid: 4633275 |
| [12] |
doi: 10.1111/j.1751-0813.2011.00695.x pmid: 21495991 |
| [13] |
doi: 10.1080/03079457.2015.1007919 |
| [14] |
pmid: 11332481 |
| [15] |
doi: 10.1292/jvms.66.1561 |
| [16] |
doi: 10.1016/j.meegid.2016.09.006 |
| [17] |
邵震, 刁有祥. 规模化鹅场主要病毒核酸检测结果与分析. 中国农业科学, 2023, 56(10): 2021-2034.
doi: 10.3864/j.issn.0578-1752.2023.10.016 |
|
doi: 10.3864/j.issn.0578-1752.2023.10.016 |
|
| [18] |
|
| [19] |
doi: 10.1080/00071668.2016.1245849 pmid: 27724044 |
| [20] |
VAN DEN BERG T P,
|
| [21] |
pmid: 24724464 |
| [22] |
doi: 10.1016/j.vetimm.2007.09.017 |
| [23] |
doi: 10.1292/jvms.14-0312 |
| [24] |
doi: S0378-1135(16)30624-1 pmid: 27938684 |
| [25] |
doi: 10.1080/03079457708418228 |
| [26] |
doi: 10.1637/11551-120516-Reg.1 pmid: 28665737 |
| [27] |
doi: 10.1080/03079457.2016.1232477 |
| [28] |
doi: 10.1111/tbed.13162 pmid: 30817083 |
| [29] |
doi: 10.1080/03079457.2016.1168513 |
| [30] |
doi: 10.1007/BF01319229 |
| [31] |
doi: 10.1016/j.vaccine.2013.12.056 pmid: 24397897 |
| [32] |
doi: 10.1637/8999-072309-Reg.1 |
| [1] | DONG Yu, WU Qian, FENG Xuan, ZHENG YinYing, CUI BaiMing. A Novel Plasmid pEA60 of Erwinia amylovora Enhances the Pathogenicity of Strains by Regulating the Synthesis of Virulence Factors [J]. Scientia Agricultura Sinica, 2026, 59(5): 996-1007. |
| [2] | CONG QiQi, ZHANG JingYi, MENG XiangLong, DAI PengBo, LI Bo, HU TongLe, WANG ShuTong, CAO KeQiang, WANG YaNan. Identification of Hypovirus in Apple Ring Rot Fungus Botryosphaeria dothidea and Detection of Virus-Carrying Status in China [J]. Scientia Agricultura Sinica, 2025, 58(3): 478-492. |
| [3] | TONG ZhaoYang, LIU WenHua, ZHANG GuoXin, DONG ChunYan, ZHANG YanXia, XU XiaoWei, HE Dong, LIU HeChun, LI Yang, WANG FengTao, FENG Jing, YAO XiaoBo, LIU MeiJin, LIN RuiMing. The Relationship Between Occurrence of Hulless Barley Ear Rot and Population Migration of Grass Mite (Siteroptes spp.) [J]. Scientia Agricultura Sinica, 2025, 58(3): 493-506. |
| [4] | YANG WenJuan, GAO JiaCheng, WANG YanTing, LI Yan, GUO Ming, WANG JunCheng, MENG YaXiong, WANG HuaJun, SI ErJing. Function of Effector Pg00778 Regulation on the Pathogenicity of Pyrenophora graminea to Barley [J]. Scientia Agricultura Sinica, 2025, 58(15): 3020-3035. |
| [5] | DONG ZaiFang, DING TengTeng, SHAN YiXuan, LI HongLian, CHEN LinLin, XING XiaoPing. Autophagy-Related Gene FpAtg3 Involves in Growth and Pathogenicity of Fusarium pseudograminearum [J]. Scientia Agricultura Sinica, 2024, 57(6): 1080-1090. |
| [6] | ZHANG AiHong, YANG Fei, ZHAO YuanYe, ZHAO YiHan, DI DianPing, MIAO HongQin. Pathogenicity and Epidemic Risk of Barley Yellow Striate Mosaic Virus [J]. Scientia Agricultura Sinica, 2024, 57(23): 4686-4697. |
| [7] | WANG Yuan, DU MengDan, LI ZhengGang, SHE XiaoMan, YU Lin, LAN GuoBing, DING ShanWen, HE ZiFu, TANG YaFei. Identification of Pathogen Causing Tomato White Tip and Curl Leaf Disease and Its Pathogenicity in Guangdong Province [J]. Scientia Agricultura Sinica, 2024, 57(12): 2350-2363. |
| [8] | ZHANG Jian, ZHAO BinSen, FENG Hao, HUANG LiLi. Function and Mechanism Analysis of Vm-milRN7 Regulating the Pathogenicity of Valsa mali [J]. Scientia Agricultura Sinica, 2024, 57(10): 1930-1942. |
| [9] | GAO XiaoXiao, TU LiQin, YANG Liu, LIU YaNan, GAO DanNa, SUN Feng, LI Shuo, ZHANG SongBai, JI YingHua. Construction of an Infectious Clone of Tobacco Mild Green Mosaic Virus Isolate Infecting Pepper from Jiangsu Based on Genomic Clone [J]. Scientia Agricultura Sinica, 2023, 56(8): 1494-1502. |
| [10] | GONG AnDong, LEI YinYu, WU NanNan, LIU JingRong, SONG MengGe, ZHANG YiMei, YANG Guang, YANG Peng. The Effect of 3-Oxyacyl ACP Reductase Gene FgOAR1 on the Growth, Development and Pathogenicity of Fusarium graminearum [J]. Scientia Agricultura Sinica, 2023, 56(24): 4854-4865. |
| [11] | YANG ShiMan, XU ChengZhi, XU BangFeng, WU YunPu, JIA YunHui, QIAO ChuanLing, CHEN HuaLan. Amino Acid of 225 in the HA Protein Affects the Pathogenicities of H1N1 Subtype Swine Influenza Viruses [J]. Scientia Agricultura Sinica, 2022, 55(4): 816-824. |
| [12] | HUANG JiaQuan,LI Li,WU FengNian,ZHENG Zheng,DENG XiaoLing. Proliferation of Two Types Prophage of ‘Candidatus Liberibacter asiaticus’ in Diaphorina citri and their Pathogenicity [J]. Scientia Agricultura Sinica, 2022, 55(4): 719-728. |
| [13] | ZHANG JinLong,ZHAO ZhiBo,LIU Wei,HUANG LiLi. The Function of Key T3SS Effectors in Pseudomonas syringae pv. actinidiae [J]. Scientia Agricultura Sinica, 2022, 55(3): 503-513. |
| [14] | LI ZhengGang,TANG YaFei,SHE XiaoMan,YU Lin,LAN GuoBing,HE ZiFu. Molecular Characteristics and Pathogenicity Analysis of Youcai Mosaic Virus Guangdong Isolate Infecting Radish [J]. Scientia Agricultura Sinica, 2022, 55(14): 2752-2761. |
| [15] | ZHANG ChengQi,LIAO LuLu,QI YongXia,DING KeJian,CHEN Li. Functional Analysis of the Nucleoporin Gene FgNup42 in Fusarium graminearium [J]. Scientia Agricultura Sinica, 2021, 54(9): 1894-1903. |
|
||