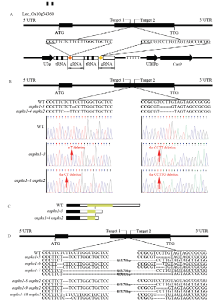
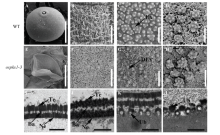
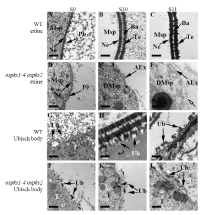
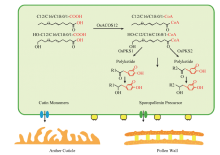
Scientia Agricultura Sinica ›› 2019, Vol. 52 ›› Issue (8): 1295-1307.doi: 10.3864/j.issn.0578-1752.2019.08.001
Special Issue: MALE STERILITY OF CROP
• MALE STERILITY OF CROP • Previous Articles Next Articles
ZHOU YuLu1,LIN Hong1,ZHANG DaBing1,2,WANG CanHua1( ),YU Jing1()
),YU Jing1()
| [1] | LONDO J P, CHIANG Y C, HUNG K H, CHIANG Y C, HUNG K H, CHIANG T Y, SCHAAL B A . Phylogeography of Asian wild rice,Oryza rufipogon, reveals multiple independent domestications of cultivated rice, Oryza sativa. Proceedings of the National Academy of Sciences of the United States of America, 2006,103(25):9578-9583. |
| [2] |
ZHANG D S, LIANG W Q, YIN C S, ZONG J, GU F W, ZHANG D B . OsC6, encoding a lipid transfer protein, is required for postmeiotic anther development in rice. Plant Physiology, 2010,154(1):149-162.
doi: 10.1104/pp.110.158865 |
| [3] |
ARIIZUMI T, TORIYAMA K . Genetic regulation of sporopollenin synthesis and pollen exine development. Annual Review of Plant Biology, 2011,62(1):437-460.
doi: 10.1146/annurev-arplant-042809-112312 |
| [4] |
JIANG J, ZHANG Z, CAO J . Pollen wall development: The associated enzymes and metabolic pathways. Plant Biology, 2013,15(2):249-263.
doi: 10.1111/plb.2013.15.issue-2 |
| [5] |
BLACKMORE S, WORTLEY H A, SKVARLA J J, ROWLEY J R . Pollen wall development in flowering plants. New Phytologist, 2007,174(3):483-498.
doi: 10.1111/nph.2007.174.issue-3 |
| [6] |
SCOTT R J, SPIELMAN M, DICKINSON H G . Stamen structure and function. The Plant Cell, 2004,16(Suppl.):S46.
doi: 10.1105/tpc.017012 |
| [7] |
SHI J X, CUI M H, YANG L, KIM Y J, ZHANG D B . Genetic and biochemical mechanisms of pollen wall development. Trends in Plant Science, 2015,20(11):741-753.
doi: 10.1016/j.tplants.2015.07.010 |
| [8] |
GOLDBERG R, BEALS T, SANDERS P . Anther development: basic principles and practical applications. The Plant Cell, 1993,5(10):1217-1229.
doi: 10.1105/tpc.5.10.1217 |
| [9] | BUBERT H, LAMBERT J, STEUERNAGEL S, AHLERS F, WIERMANN R . Continuous decomposition of sporopollenin from pollen of Typha angustifolia L. by acidic methanolysis. Zeitschrift Fur Naturforschung Section C-a Journal of Biosciences, 2002,57(11/12):1035-1041. |
| [10] | BROOKS J, SHAW G . Chemical structure of the exine of pollen walls and a new function for carotenoids in nature. Nature, 1968,219(5153):532-533. |
| [11] | SCOTT R J, STEAD A D. Molecular and Cellular Aspects of Plant Reproduction. England: Cambridge University Press, 1994: 49-81. |
| [12] |
HUI L, PINOT F, SAUVEPLANE V, WERCK-REICHHART D, DIEHL P, SCHREIBER L, FRANKE R, ZHANG P, CHEN L, GAO Y W, LIANG W Q, ZHANG D B . Cytochrome P450 family member CYP704B2 catalyzes the {omega-hydroxylation of fatty acids and is required for anther cutin biosynthesis and pollen exine formation in rice. The Plant Cell, 2010,22(9):173-90.
doi: 10.1105/tpc.109.070326 |
| [13] |
YANG X J, WU D, SHI J X, HE Y, PINOT F, GRAUSEM B, YIN C S, ZHU L, CHEN M J, LUO Z J, LIANG W Q, ZHANG D B . Rice CYP703A3, a cytochrome P450 hydroxylase, is essential for development of anther cuticle and pollen exine. Journal of Integrative Plant Biology, 2014,56(10):979-994.
doi: 10.1111/jipb.v56.10 |
| [14] | DOBRITSA A A, SHRESTHA J, MORANT M, PINOT F, MATSUNO M, SWANSON R, MØLLER B L, PREUSS D . CYP704B1 is a long-chain fatty acid ω-hydroxylase essential for sporopollenin synthesis in pollen of Arabidopsis. Plant Physiology, 2009,151(2):574-589. |
| [15] |
MORANT M, JØRGENSEN K, SCHALLER H, PINOT F, MØLLER B L, WERCK-REICHHART D, BAKA S . CYP703 is an ancient cytochrome P450 in land plants catalyzing in-chain hydroxylation of lauric acid to provide building blocks for sporopollenin synthesis in pollen. The Plant Cell, 2007,19(5):1473-1487.
doi: 10.1105/tpc.106.045948 |
| [16] |
SHI J, TAN H X, YU X H, LIU Y Y, LIANG W Q, RANATHUNGE K, FRANKE R B, SCHREIBER L, WANG Y J, KAI G Y, SHANKLIN J, MA H, ZHANG D B . Defective Pollen Wall is required for anther and microspore development in rice and encodes a fatty acyl carrier protein reductase. The Plant Cell, 2011,23(6):2225-2246.
doi: 10.1105/tpc.111.087528 |
| [17] | CHEN W W, YU X H, ZHANG K S, SHI J X, OLIVEIRA S D, SCHREIBER L, SHANKLIN J, ZHANG D B . Male Sterile2 encodes a plastid-localized fatty acyl carrier protein reductase required for pollen exine development in Arabidopsis. Plant Physiology, 2011,157(2):842-853. |
| [18] | XU D W, SHI J X, RAUTENGARTEN C, YANG L, QIAN X L, UZAIR M, ZHU L, LUO Q, AN G H, WAßMANN F, SCHREIBER L, HEAZLEWOOD J L, SCHELLER H V, HU J P, ZHANG D B, LIANG W Q . Defective Pollen Wall 2 (DPW2) encodes an acyl transferase required for rice pollen development. Plant Physiology, 2016,173(1):240-255. |
| [19] |
LIU Z, LIN S, SHI J X, YU J, ZHU L, YANG X J, ZHANG D B, LIANG W Q . Rice No Pollen 1 (NP1) is required for anther cuticle formation and pollen exine patterning. The Plant Journal, 2017,91(2):263-277.
doi: 10.1111/tpj.2017.91.issue-2 |
| [20] |
JEPSON C, KARPPINEN K, DAKU R M, STERENBERG B T, SUH D Y , . Hypericum perforatum hydroxyalkylpyrone synthase involved in sporopollenin biosynthesis--phylogeny, site-directed mutagenesis, and expression in nonanther tissues. The FEBS Journal, 2014,281(17):3855-3868.
doi: 10.1111/febs.2014.281.issue-17 |
| [21] | LI Y L, LI D D, GUO Z L, SHI Q S, XIONG S X, ZHANG C, ZHU J, YANG Z N . OsACOS12, an orthologue of Arabidopsis acyl-CoA synthetase5, plays an important role in pollen exine formation and anther development in rice. BMC Plant Biology, 2016,16(1):256. |
| [22] | QIN M M, TIAN T T, XIA S Q, WANG Z X, SONG L P Y B, QIN WEN J, SHEN J X, MA C Z, FU T D, TU J X . Heterodimer formation of BnPKSA or BnPKSB with BnACOS5 constitutes a multienzyme complex in tapetal cells and is involved in male reproductive development in Brassica napus. Plant and Cell Physiology, 2016,57(8):1643-1656. |
| [23] |
YANG X J, LIANG W Q, CHEN M J, ZHANG D B, ZHAO X X, SHI J X . Rice fatty acyl-CoA synthetase OsACOS12 is required for tapetum programmed cell death and male fertility. Planta, 2017,246(1):105-122.
doi: 10.1007/s00425-017-2691-y |
| [24] |
WANG Y B, LIN Y C, SO J, DU Y G, LO C . Conserved metabolic steps for sporopollenin precursor formation in tobacco and rice. Physiologia Plantarum, 2013,149(1):13-24.
doi: 10.1111/ppl.2013.149.issue-1 |
| [25] | SOUZA C D A, KIM S S, KOCH S, KIENOW L, SCHNEIDER K, MCKIM S M, HAUGHN G W, KOMBRINK E, DOUGLAS C J . A novel fatty Acyl-CoA Synthetase is required for pollen development and sporopollenin biosynthesis in Arabidopsis. The Plant Cell, 2009,21(2):507-525. |
| [26] | XIE L L, LIU P L, ZHU Z X, ZHANG S F, ZHANG S J, LI F, ZHANG H, LI G L, WEI Y X, SUN R F . Phylogeny and expression analyses reveal important roles for plant PKS III family during the conquest of land by plants and angiosperm diversification. Frontiers in Plant Science, 2016,7(136):1312. |
| [27] | KIM S S, GRIENENBERGER E, LALLEMAND B, COLPITTS C C, KIM S Y, SOUZA C D A, GEOFFROY P, HEINTZ D, KRAHN D, KAISER M, KOMBRINK E, HEITZ T, SUH D Y, LEGRAND M, DOUGLASA C J . LAP6/POLYKETIDE SYNTHASE a and LAP5/POLYKETIDE SYNTHASE B encode hydroxyalkyl α-pyrone synthases required for pollen development and sporopollenin biosynthesis in Arabidopsis thaliana. The Plant Cell, 2010,22(12):4045-4066. |
| [28] | GRIENENBERGER E, KIM S S, LALLEMAND B, GEOFFROY P, HEINTZ D, SOUZA C D A, HEITZ T, DOUGLAS C J, LEGRAND M . Analysis of TETRAKETIDE α-PYRONE REDUCTASE function in Arabidopsis thaliana reveals a previously unknown, but conserved, biochemical pathway in sporopollenin monomer biosynthesis. The Plant Cell, 2010,22(12):4067-4083. |
| [29] |
ZHU X L, YU J, SHI J X, TOHGE T, FERNIE A R, MEIR S, AHARONI A, XU D W, ZHANG D B, LIANG W Q . The polyketide synthase OsPKS2 is essential for pollen exine and Ubisch body patterning in rice. Journal of Integrative Plant Biology, 2017,59(9):612-628.
doi: 10.1111/jipb.12574 |
| [30] | SHI Q S, WANG K Q, LI Y L, ZHOU L, XIONG S X, HAN Y, ZHANG Y F, YANG N Y, YANG Z N, ZHU J . OsPKS1 is required for sexine layer formation, which shows functional conservation between rice and Arabidopsis. Plant Science, 2018,277:145-154. |
| [31] | DOBRITSA A A, LEI Z T, NISHIKAWA S I, URBANCZYK- WOCHNIAK E, HUHMAN D V, PREUSS D, SUMNER L W . LAP5 and LAP6 encode anther-specific proteins with similarity to chalcone synthase essential for pollen exine development in Arabidopsis. Plant Physiology, 2010,153(3):937-955. |
| [32] |
ZOU T, XIAO Q, LI W J, LUO T, YUAN G Q, HE Z Y, LIU M X, LI Q, XU P Z, ZHU J, LIANG Y Y, DENG Q M, WANG S Q, ZHENG A P, WANG L X, LI P and LI S C . OsLAP6/OsPKS1, an orthologue of Arabidopsis PKSA/LAP6, is critical for proper pollen exine formation. Rice, 2017,10(1):53.
doi: 10.1186/s12284-017-0191-0 |
| [33] |
XIE K B, MINKENBERG B, YANG Y N . Boosting CRISPR/Cas9 multiplex editing capability with the endogenous tRNA-processing system. Proceedings of the National Academy of Sciences of the USA, 2015,112(11):3570-3575.
doi: 10.1073/pnas.1420294112 |
| [34] |
LI N, ZHANG D S, LIU H S, YIN C S, LI X X, LIANG W Q, YUAN Z, XU B, CHU H W, WANG J, WEN T Q, HUANG H, LUO D, MA H, ZHANG D B . The rice tapetum degeneration retardation gene is required for tapetum degradation and anther development. The Plant Cell, 2006,18(11):2999-3014.
doi: 10.1105/tpc.106.044107 |
| [35] |
ZHANG D B, WILSON Z A . Stamen specification and anther development in rice. Chinese Science Bulletin, 2009,54(14):2342-2353.
doi: 10.1007/s11434-009-0348-3 |
| [36] |
ZHANG D B, LUO X, ZHU L . Cytological analysis and genetic control of rice anther development. Journal of Genetics and Genomics, 2011,38(9):379-390.
doi: 10.1016/j.jgg.2011.08.001 |
| [37] |
LIN H, YU J, PEARCE S P, ZHANG D B, WILSON Z A . RiceAntherNet: A gene co-expression network for identifying anther and pollen development genes. The Plant Journal, 2017,92(6):1076-1091.
doi: 10.1111/tpj.13744 |
| [38] | MEN X, SHI J X, LIANG W Q, ZHANG Q F, LIAN G B, QUAN S, ZHU L, LUO Z J, CHEN M J, ZHANG D B . Glycerol-3-Phosphate Acyltransferase 3 (OsGPAT3) is required for anther development and male fertility in rice. Journal of Experimental Botany, 2017,68(3):513-526. |
| [1] | ZHAO ChunFang,ZHAO QingYong,LÜ YuanDa,CHEN Tao,YAO Shu,ZHAO Ling,ZHOU LiHui,LIANG WenHua,ZHU Zhen,WANG CaiLin,ZHANG YaDong. Screening of Core Markers and Construction of DNA Fingerprints of Semi-Waxy Japonica Rice Varieties [J]. Scientia Agricultura Sinica, 2022, 55(23): 4567-4582. |
| [2] | PANG HongBo, CHENG Lu, YU MingLan, CHEN Qiang, LI YueYing, WU LongKun, WANG Ze, PAN XiaoWu, ZHENG XiaoMing. Genome-Wide Association Study of Cold Tolerance at the Germination Stage of Rice [J]. Scientia Agricultura Sinica, 2022, 55(21): 4091-4103. |
| [3] | DENG AiXing,LIU YouHong,MENG Ying,CHEN ChangQing,DONG WenJun,LI GeXing,ZHANG Jun,ZHANG WeiJian. Effects of 1.5℃ Field Warming on Rice Yield and Quality in High Latitude Planting Area [J]. Scientia Agricultura Sinica, 2022, 55(1): 51-60. |
| [4] | YinHua MA,KaiQin MO,Lu LIU,PingFang LI,ChenZhong JIN,Fang YANG. Effect of Overexpression of OsRRK1 Gene on Rice Leaf Development [J]. Scientia Agricultura Sinica, 2021, 54(5): 877-886. |
| [5] | ZHANG YaDong,LIANG WenHua,HE Lei,ZHAO ChunFang,ZHU Zhen,CHEN Tao,ZHAO QingYong,ZHAO Ling,YAO Shu,ZHOU LiHui,LU Kai,WANG CaiLin. Construction of High-Density Genetic Map and QTL Analysis of Grain Shape in Rice RIL Population [J]. Scientia Agricultura Sinica, 2021, 54(24): 5163-5176. |
| [6] | XU ZiYi,CHENG Xing,SHEN Qi,ZHAO YaNan,TANG JiaYu,LIU Xi. Identification and Gene Functional Analysis of Yellow Green Leaf Mutant ygl3 in Rice [J]. Scientia Agricultura Sinica, 2021, 54(15): 3149-3157. |
| [7] | REN ZhiJie,LI Qian,SUN YuJia,KONG DongDong,LIU LiangYu,HOU CongCong,LI LeGong. OsCSC11 Mediates Dry-Hot Wind/Drought-Induced Ca2+ Signal to Regulate Stamen Development in Rice [J]. Scientia Agricultura Sinica, 2021, 54(10): 2039-2052. |
| [8] | LIU HaiYing,FENG BiDe,RU ZhenGang,CHEN XiangDong,HUANG PeiXin,XING ChenTao,PAN YinYin,ZHEN JunQi. Relationship Between Phytohormones and Male Sterility of BNS and BNS366 in Wheat [J]. Scientia Agricultura Sinica, 2021, 54(1): 1-18. |
| [9] | KunNeng ZHOU,JiaFa XIA,Peng YUN,YuanLei WANG,TingChen MA,CaiJuan ZHANG,ZeFu LI. Transcriptome Research of Erect and Short Panicle Mutant esp in Rice [J]. Scientia Agricultura Sinica, 2020, 53(6): 1081-1094. |
| [10] | ShuJun MENG,XueHai ZHANG,QiYue WANG,Wen ZHANG,Li HUANG,Dong DING,JiHua TANG. Identification of miRNAs and tRFs in Response to Salt Stress in Rice Roots [J]. Scientia Agricultura Sinica, 2020, 53(4): 669-682. |
| [11] | PAN ZhengYan,LIU Bo,JIANG HongBo,YAO JiPan,BAI YuanJun,XU ZhengJin. Effect of Panicle Neck Blast on Grain Yield and Stem Node Metabolites at the Rice Filling Stage [J]. Scientia Agricultura Sinica, 2020, 53(20): 4177-4188. |
| [12] | WANG FangQuan,CHEN ZhiHui,XU Yang,WANG Jun,LI WenQi,FAN FangJun,CHEN LiQin,TAO YaJun,ZHONG WeiGong,YANG Jie. Development and Application of the Functional Marker for the Broad-Spectrum Blast Resistance Gene PigmR in Rice [J]. Scientia Agricultura Sinica, 2019, 52(6): 955-967. |
| [13] | ZHOU JiaQin,ZHU JunZhao,YANG SiXue,ZHU ZhouJie,YAO Jie,ZHENG WenJuan,ZHU ShiHua,DING WoNa. Cloning and Functional Analysis of a Root Development Related Gene OsKSR7 in Rice (Oryza sativa L.) [J]. Scientia Agricultura Sinica, 2019, 52(5): 777-785. |
| [14] | Mu ZHANG, ShuanHu TANG, QiaoYi HUANG, YuWan PANG, Qiong YI, Xu HUANG, Ping LI, HongTing FU. The Nutrient Supply Characteristics of Co-Application of Slow-Release Urea and Common Urea in Double-Cropping Rice [J]. Scientia Agricultura Sinica, 2018, 51(20): 3985-3995. |
| [15] | TAO XingLin, HOU Dong, ZHU HuiXia, LIU MingXia, ZHANG JinWen, HU LiMin. Transcriptome and Cytological Researches on the Anther Abortion of a Thermo-Sensitive Genic Male Sterile Line GS-19 in Cauliflower [J]. Scientia Agricultura Sinica, 2017, 50(13): 2538-2552. |
|
||