






中国农业科学 ›› 2021, Vol. 54 ›› Issue (8): 1599-1612.doi: 10.3864/j.issn.0578-1752.2021.08.003
孟鑫浩1( ),邓洪涛1(),李丽2,崔顺立1,Charles Y.Chen3,侯名语1,杨鑫雷1(),刘立峰1()
),邓洪涛1(),李丽2,崔顺立1,Charles Y.Chen3,侯名语1,杨鑫雷1(),刘立峰1()
收稿日期:2020-09-23
接受日期:2020-12-07
出版日期:2021-04-16
发布日期:2021-04-25
通讯作者:
杨鑫雷,刘立峰
作者简介:孟鑫浩,E-mail: 基金资助:
MENG XinHao1(),DENG HongTao1(),LI Li2,CUI ShunLi1,Charles Y. CHEN3,HOU MingYu1,YANG XinLei1(),LIU LiFeng1()
Received:2020-09-23
Accepted:2020-12-07
Online:2021-04-16
Published:2021-04-25
Contact:
XinLei YANG,LiFeng LIU
摘要:
【目的】栽培种花生是世界范围内重要的油料作物和经济作物,其株型相关性状是典型的数量性状,亦是重要的农艺性状,与产量和机械化收获密切相关。对花生株型相关性状进行遗传分析和QTL定位,筛选与之紧密连锁的分子标记,有助于花生的品种保护和品种鉴别,为花生株型分子育种提供重要的理论依据。【方法】以直立型花生品种冀花5号和匍匐型M130为亲本构建的包含321个家系的RIL群体为研究材料,于2016—2018年分别在海南市、邯郸市、保定市和唐山市等7个环境下种植,各个环境均在收获时调查统计花生侧枝夹角、主茎高、侧枝长、株型指数和扩展半径等5个株型相关性状的表型值。同时,利用SSR、AhTE、SRAP和TRAP等分子标记扫描亲本及群体的基因型用于构建分子遗传连锁图谱。最后结合多年多点的表型数据,采用QTL Icimapping V4.2中的完备区间作图法(inclusive composite interval mapping,ICIM)对7个环境下的株型相关性状进行加性QTL和上位性QTL分析。【结果】构建了一张包含363个多态性位点的分子遗传连锁图谱,所有标记被分配到20条染色体和1个未知连锁群;图谱总长度覆盖全基因组的1 360.38 cM,标记间平均距离为3.75 cM;单个连锁群长度为39.599—101.056 cM,包括4—50个分子标记。共检测到30个与株型相关性状的加性QTL,分布在A04、A05、A06、A08、A09、B02、B09等7条染色体上。其中,5个QTL与侧枝夹角相关,可解释的表型变异(phenotypic variance explained,PVE)为3.48%—11.22%;15个QTL与主茎高相关,PVE为3.58%—10.05%;2个QTL与侧枝长相关,PVE为6.03%—8.56%;4个QTL与株型指数相关,PVE为4.68%—15.08%;4个QTL与扩展半径相关,PVE为3.30%—9.33%。qLBAA05.1、qLBAA05.2、qMSHA04.2和qIOPTA05.1等4个主效QTL,可解释的表型变异分别为11.22%、10.59%、10.23%、10.05%和15.08%。此外,共检测到59对上位性QTL。其中,6对上位性QTL与侧枝长相关,PVE为2.23%—2.78%;50对上位性QTL与株型指数相关,PVE为0.25%—1.44%;3对上位性QTL与扩展半径相关,PVE为7.28%—12.25%。发现3个QTL聚集区,分别为A04染色体的GM1867—AHGS1967区间、A05染色体的me14em5-116—PM418区间和A08染色体的HBAUAh177—AhTE0658区间,涉及侧枝夹角、主茎高、株型指数和扩展半径等4个株型相关性状。【结论】构建了一张包含363个标记位点的分子遗传连锁图谱;获得30个与株型相关性状的加性QTL和59对上位性QTL;发现3个QTL聚集区。
孟鑫浩,邓洪涛,李丽,崔顺立,Charles Y.Chen,侯名语,杨鑫雷,刘立峰. 栽培种花生株型相关性状的QTL定位[J]. 中国农业科学, 2021, 54(8): 1599-1612.
MENG XinHao,DENG HongTao,LI Li,CUI ShunLi,Charles Y. CHEN,HOU MingYu,YANG XinLei,LIU LiFeng. QTL Mapping for Lateral Branch Angle Related Traits of Cultivated Peanut (Arachis hypogaea L.)[J]. Scientia Agricultura Sinica, 2021, 54(8): 1599-1612.
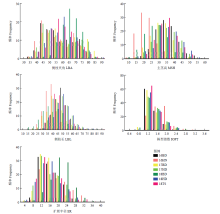
图1
7个环境下RIL群体与亲本的5个性状的正态检验和频率分布图 环境16BD、16HN、17BD、17HD、18BD、18HD和18TS分别代表2016年、2017年和2018年保定市、海南市、邯郸市和唐山市;LBA:侧枝夹角;MSH:主茎高;LBL:侧枝长;IOPT:株型指数;ER:扩展半径"
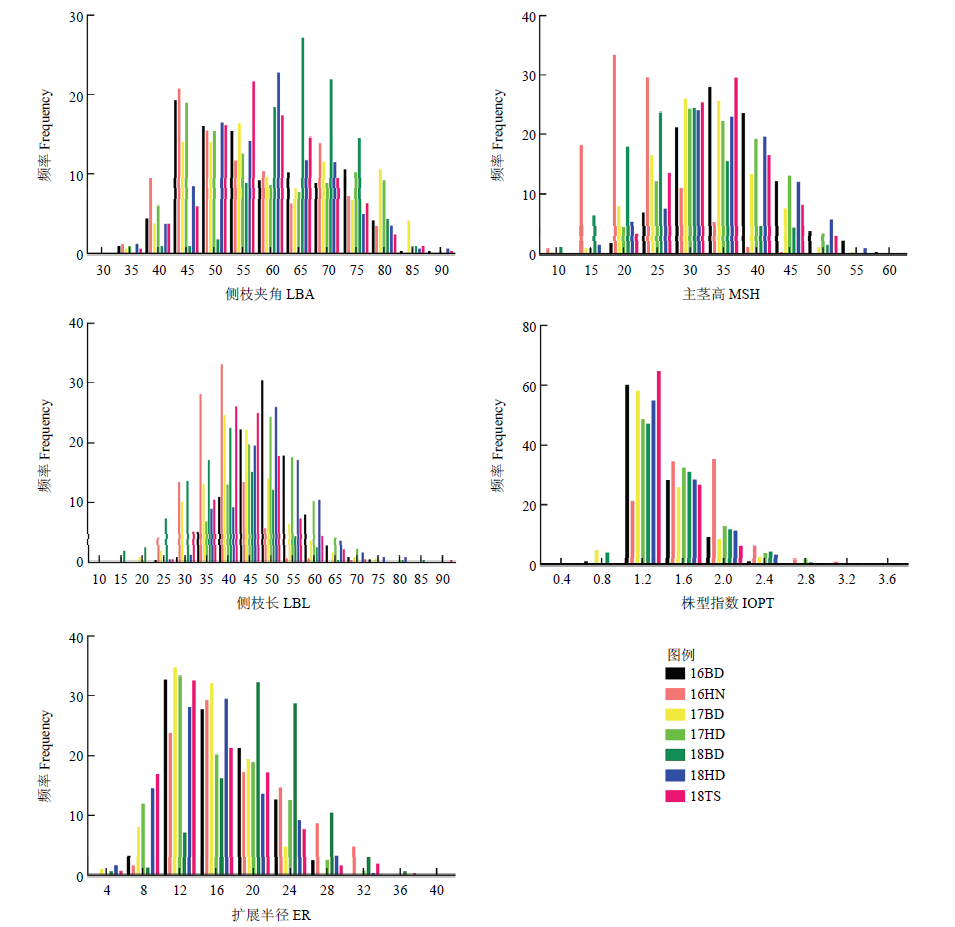
表1
亲本及RIL群体表型数据统计分析"
| 性状 Trait | 环境 Env. | 亲本 Parent | RIL群体RILs population | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 冀花5号 Jihua 5 | M130 | 最小值 Minimum | 最大值 Maximum | 平均值 Mean | 标准差 SD | 变异系数 CV (%) | Shapiro- wilk (w) | 偏度 Skewness | 峰度 Kurtosis | ||
| 侧枝夹角 LBA (°) | 16BD | 45.93 | 81.22 | 34.15 | 87.08 | 57.61 | 11.64 | 20.20 | 0.955*** | 0.330 | -0.993 |
| 16HN | 37.41 | 77.05 | 34.03 | 80.07 | 55.90 | 11.75 | 21.02 | 0.953*** | 0.362 | -1.094 | |
| 17BD | 43.50 | 83.43 | 33.07 | 87.52 | 60.47 | 12.80 | 21.17 | 0.964*** | 0.277 | -1.050 | |
| 17HD | 39.20 | 79.01 | 35.35 | 90.10 | 58.63 | 13.09 | 22.33 | 0.949*** | 0.344 | -1.128 | |
| 18BD | 37.23 | 72.47 | 39.01 | 85.73 | 65.93 | 7.64 | 11.58 | 0.996 | -0.344 | 0.408 | |
| 18HD | 45.30 | 81.83 | 35.05 | 88.38 | 58.78 | 10.53 | 17.91 | 0.989 | 0.252 | -0.246 | |
| 18TS | 47.50 | 78.73 | 36.05 | 89.00 | 58.93 | 9.87 | 16.76 | 0.989 | 0.327 | -0.136 | |
| 主茎高 MSH (cm) | 16BD | 45.05 | 23.71 | 19.10 | 59.57 | 36.33 | 6.97 | 19.18 | 0.988** | 0.348 | 0.221 |
| 16HN | 35.05 | 17.31 | 10.36 | 45.00 | 22.67 | 5.82 | 25.66 | 0.969*** | 0.702 | 0.415 | |
| 17BD | 38.40 | 19.37 | 10.15 | 56.62 | 32.25 | 7.17 | 22.25 | 0.996 | 0.120 | -0.038 | |
| 17HD | 45.26 | 25.52 | 13.37 | 52.34 | 34.30 | 7.62 | 22.22 | 0.991 | -0.039 | -0.481 | |
| 18BD | 38.00 | 19.70 | 10.71 | 52.52 | 27.96 | 7.94 | 28.38 | 0.977** | 0.475 | 0.164 | |
| 18HD | 45.05 | 23.36 | 12.28 | 55.76 | 34.89 | 8.04 | 23.04 | 0.992 | -0.102 | -0.087 | |
| 18TS | 45.23 | 24.38 | 17.06 | 51.10 | 33.87 | 6.69 | 19.74 | 0.99 | 0.161 | -0.297 | |
| 侧枝长 LBL (cm) | 16BD | 49.76 | 61.42 | 24.56 | 75.71 | 49.20 | 7.56 | 15.36 | 0.995 | 0.144 | 0.642 |
| 16HN | 38.81 | 36.26 | 23.52 | 70.33 | 38.13 | 6.51 | 17.08 | 0.966*** | 0.689 | 2.532 | |
| 17BD | 40.36 | 46.26 | 19.43 | 76.87 | 42.85 | 9.30 | 21.69 | 0.991 | 0.534 | 0.954 | |
| 17HD | 47.68 | 55.30 | 23.00 | 83.00 | 49.71 | 8.74 | 17.59 | 0.979** | 0.409 | 0.576 | |
| 18BD | 41.00 | 76.71 | 14.27 | 87.81 | 39.25 | 10.13 | 25.80 | 0.996 | 0.462 | 1.660 | |
| 18HD | 49.76 | 61.38 | 25.52 | 79.31 | 49.52 | 9.01 | 18.19 | 0.988 | 0.316 | 0.623 | |
| 18TS | 51.53 | 61.67 | 24.05 | 90.00 | 44.83 | 8.29 | 18.50 | 0.955*** | 0.813 | 2.377 | |
| 株型指数 IOPT | 16BD | 1.02 | 2.76 | 0.95 | 2.75 | 1.39 | 0.27 | 19.67 | 0.932*** | 1.174 | 1.470 |
| 16HN | 1.01 | 2.21 | 1.05 | 3.27 | 1.73 | 0.38 | 21.80 | 0.958*** | 0.892 | 1.533 | |
| 17BD | 1.05 | 2.23 | 0.76 | 2.86 | 1.37 | 0.34 | 24.63 | 0.93*** | 1.197 | 1.632 | |
| 17HD | 1.03 | 2.29 | 1.01 | 3.02 | 1.51 | 0.39 | 25.96 | 0.879*** | 1.479 | 2.665 | |
| 18BD | 1.07 | 3.79 | 0.68 | 3.45 | 1.46 | 0.40 | 27.27 | 0.933*** | 1.297 | 2.645 | |
| 18HD | 1.05 | 2.76 | 1.02 | 3.51 | 1.48 | 0.39 | 26.41 | 0.829*** | 1.961 | 5.759 | |
| 18TS | 1.15 | 2.42 | 1.06 | 2.57 | 1.34 | 0.29 | 21.41 | 0.917*** | 0.654 | 3.582 | |
| 扩展半径 ER (cm) | 16BD | 8.52 | 22.35 | 7.75 | 27.06 | 16.50 | 4.49 | 27.20 | 0.974*** | 0.411 | -0.747 |
| 16HN | 16.13 | 31.57 | 8.05 | 33.08 | 18.52 | 5.80 | 31.34 | 0.965*** | 0.593 | -0.562 | |
| 17BD | 9.37 | 21.30 | 5.02 | 25.06 | 15.05 | 3.98 | 26.46 | 0.99 | 0.181 | -0.517 | |
| 17HD | 10.29 | 19.39 | 6.51 | 33.05 | 15.89 | 5.32 | 33.47 | 0.971*** | 0.52 | -0.451 | |
| 18BD | 9.33 | 34.71 | 5.38 | 36.38 | 21.00 | 5.01 | 23.83 | 0.996 | -0.035 | 0.345 | |
| 18HD | 8.67 | 21.17 | 5.01 | 30.02 | 15.11 | 5.16 | 34.15 | 0.978** | 0.477 | -0.283 | |
| 18TS | 9.50 | 24.13 | 5.71 | 34.05 | 15.07 | 5.49 | 36.44 | 0.961*** | 0.742 | 0.357 | |
表2
栽培种花生株型相关性状的相关分析"
| 环境Env. | 性状Trait | 侧枝夹角LBA | 主茎高MSH | 侧枝长LBL | 株型指数IOPT |
|---|---|---|---|---|---|
| 16BD | 主茎高MSH | -0.323** | |||
| 侧枝长LBL | 0.323** | 0.456** | |||
| 株型指数IOPT | 0.604** | -0.631** | 0.374** | ||
| 扩展半径ER | 0.820** | -0.262** | 0.450** | 0.646** | |
| 16HN | 主茎高MSH | -0.127* | |||
| 侧枝长LBL | -0.009* | 0.584** | |||
| 株型指数IOPT | 0.205** | -0.706** | 0.061* | ||
| 扩展半径ER | 0.761** | 0.086* | 0.223** | 0.129* | |
| 17BD | 主茎高MSH | -0.278** | |||
| 侧枝长LBL | 0.429** | 0.450** | |||
| 株型指数IOPT | 0.683** | -0.539** | 0.473** | ||
| 扩展半径ER | 0.736** | -0.093* | 0.589** | 0.651** | |
| 17HD | 主茎高MSH | -0.372** | |||
| 侧枝长LBL | 0.333** | 0.394** | |||
| 株型指数IOPT | 0.595** | -0.687** | 0.349** | ||
| 扩展半径ER | 0.820** | -0.270** | 0.496** | 0.638** | |
| 18BD | 主茎高MSH | -0.118* | |||
| 侧枝长LBL | 0.557** | 0.547** | |||
| 株型指数IOPT | 0.457** | -0.506** | 0.408** | ||
| 扩展半径ER | 0.815** | 0.303** | 0.748** | 0.455** | |
| 18HD | 主茎高MSH | -0.184** | |||
| 侧枝长LBL | 0.458** | 0.466** | |||
| 株型指数IOPT | 0.537** | -0.666** | 0.281** | ||
| 扩展半径ER | 0.897** | -0.155** | 0.546** | 0.577** | |
| 18TS | 主茎高MSH | -0.117* | |||
| 侧枝长LBL | 0.380** | 0.538** | |||
| 株型指数IOPT | 0.465** | -0.532** | 0.352** | ||
| 扩展半径ER | 0.878** | -0.044 | 0.512** | 0.518** |
表3
RIL群体各性状方差分析及广义遗传力"
| 性状 Trait | 变异来源 Variables | 自由度 df | 均方 MS | F值 F-value | P值 P-value | 广义遗传力 h2B |
|---|---|---|---|---|---|---|
| 侧枝夹角 LBA | 基因型Genotype | 320 | 616.890 | 14.658 | <0.01 | 0.86 |
| 环境Environment | 6 | 3124.760 | 74.247 | <0.01 | ||
| 基因型×环境G×E | 1877 | 42.086 | 1.036 | >0.05 | ||
| 主茎高 MSH | 基因型Genotype | 320 | 240.144 | 11.943 | <0.01 | 0.92 |
| 环境Environment | 6 | 7341.320 | 365.118 | <0.01 | ||
| 基因型×环境G×E | 1877 | 20.107 | 1.723 | <0.01 | ||
| 侧枝长 LBL | 基因型Genotype | 320 | 270.164 | 6.924 | <0.01 | 0.90 |
| 环境Environment | 6 | 7396.938 | 189.580 | <0.01 | ||
| 基因型×环境G×E | 1779 | 39.017 | 1.836 | <0.01 | ||
| 株型指数 IOPT | 基因型Genotype | 320 | 0.466 | 7.433 | <0.01 | 0.91 |
| 环境Environment | 6 | 4.097 | 65.320 | <0.01 | ||
| 基因型×环境G×E | 1779 | 0.063 | 1.896 | <0.01 | ||
| 扩展半径 ER | 基因型Genotype | 320 | 121.227 | 12.600 | <0.01 | 0.89 |
| 环境Environment | 6 | 1599.042 | 166.201 | <0.01 | ||
| 基因型×环境G×E | 1881 | 9.621 | 2.265 | <0.01 |
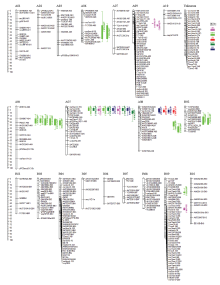
图2
栽培种花生遗传连锁图谱"

表4
株型相关性状的QTL定位结果"
| 性状 Trait | QTL | 环境 Env. | 染色体 Chr. | 位置 Position | 标记区间 Marker interval | LOD | 贡献率 PVE (%) | 加性效应 Additive |
|---|---|---|---|---|---|---|---|---|
| 侧枝夹角 LBA | qLBAA05.1 | 16BD | A05 | 6.00 | me14em5-116—PM418 | 7.44 | 11.22 | -3.9527 |
| 17BD | A05 | 6.00 | me14em5-116—PM418 | 6.99 | 10.59 | -4.2307 | ||
| 18HD | A05 | 6.00 | me14em5-116—PM418 | 6.37 | 8.17 | -3.2618 | ||
| qLBAA05.2 | 16HN | A05 | 5.00 | me14em5-116—PM418 | 5.37 | 8.62 | -3.6198 | |
| 17HD | A05 | 5.00 | me14em5-116—PM418 | 7.22 | 10.23 | -4.8094 | ||
| qLBAA09.1 | 18BD | A09 | 20.98 | GM66—GM1076 | 3.13 | 4.34 | -2.0477 | |
| qLBAA09.2 | 16HN | A09 | 24.98 | RN27A10—AHTE0122 | 2.91 | 3.48 | -2.3139 | |
| qLBAB09.1 | 18HD | B09 | 50.00 | me5em5-100—me13em8-142 | 2.89 | 4.51 | 2.4198 | |
| 主茎高 MSH | qMSHA04.1 | 17BD | A04 | 21.00 | GM1867—AHGS1967 | 4.94 | 6.23 | 1.9109 |
| qMSHA04.2 | 18BD | A04 | 22.00 | GM1867—AHGS1967 | 7.49 | 10.05 | 2.7157 | |
| qMSHA04.3 | 18HD | A04 | 23.00 | GM1867—AHGS1967 | 3.55 | 5.22 | 1.9577 | |
| qMSHA04.4 | 16BD | A04 | 24.00 | GM1867—AHGS1967 | 3.11 | 3.58 | 1.6317 | |
| qMSHA04.5 | 16HN | A04 | 26.00 | GM1867—AHGS1967 | 3.99 | 5.24 | 1.4725 | |
| qMSHA04.6 | 17HD | A04 | 28.00 | GM1867—AHGS1967 | 6.54 | 6.34 | 2.4193 | |
| qMSHA05.1 | 16BD | A05 | 4.00 | me14em5-116 - PM418 | 4.54 | 6.43 | 2.1285 | |
| qMSHA05.2 | 17HD | A05 | 5.00 | me14em5-116—PM418 | 4.73 | 5.26 | 2.1268 | |
| qMSHA06.1 | 16HN | A06 | 36.41 | TC7C06—AHTE0372 | 4.54 | 4.90 | 1.3824 | |
| 17BD | A06 | 36.41 | TC7C06—AHTE0372 | 3.46 | 4.23 | 1.5366 | ||
| qMSHA06.2 | 18BD | A06 | 46.21 | TC7C06—AHTE0372 | 3.21 | 3.67 | 1.5995 | |
| qMSHA08.1 | 17BD | A08 | 30.00 | Ah4-4—TC9B08 | 3.58 | 4.62 | -1.6048 | |
| qMSHA08.2 | 18BD | A08 | 35.00 | TC9B08—AHGS1947b | 3.36 | 4.47 | -1.7650 | |
| qMSHB02.1 | 17HD | B02 | 25.00 | AHTE0398—CTW_NEW_38 | 4.00 | 4.48 | -1.9683 | |
| qMSHB02.2 | 18HD | B02 | 29.00 | AHTE0398—CTW_NEW_38 | 2.96 | 3.74 | -1.6183 | |
| qMSHB09.1 | 16HN | B09 | 22.00 | AHGS1576—me11em4-144 | 7.92 | 9.44 | -1.9154 | |
| 侧枝长 LBL | qLBLA08.1 | 18BD | A08 | 8.40 | HBAUAh177—AhTE0658 | 4.86 | 6.03 | -3.2587 |
| qLBLA08.2 | 17BD | A08 | 9.12 | HBAUAh177—AhTE0658 | 5.27 | 7.53 | -2.5945 | |
| 18HD | A08 | 9.12 | HBAUAh177—AhTE0658 | 6.13 | 8.56 | -2.6868 | ||
| 株型指数 IOPT | qIOPTA05.1 | 16BD | A05 | 4.00 | me14em5-116—PM418 | 10.15 | 15.08 | -0.1151 |
| 17HD | A05 | 4.00 | me14em5-116—PM418 | 5.18 | 9.35 | -0.1308 | ||
| qIOPTA05.2 | 18TS | A05 | 5.00 | me14em5-116—PM418 | 3.84 | 5.85 | -0.0757 | |
| qIOPTA05.3 | 17BD | A05 | 6.00 | me14em5-116—PM418 | 6.25 | 8.58 | -0.1006 | |
| 18BD | A05 | 6.00 | me14em5-116—PM418 | 3.23 | 4.68 | -0.1121 | ||
| qIOPTA05.4 | 18HD | A05 | 7.00 | PM418—pPGSSeq11E11 | 4.48 | 6.47 | -0.1012 | |
| 扩展半径 ER | qERA05.1 | 16BD | A05 | 5.00 | me14em5-116—PM418 | 6.63 | 9.33 | -1.5728 |
| 16HN | A05 | 5.00 | me14em5-116—PM418 | 5.03 | 7.41 | -1.7923 | ||
| 17BD | A05 | 5.00 | me14em5-116—PM418 | 3.16 | 3.86 | -0.9281 | ||
| 17HD | A05 | 5.00 | me14em5-116—PM418 | 6.15 | 8.55 | -1.8270 | ||
| 18HD | A05 | 5.00 | me14em5-116—PM418 | 5.14 | 7.38 | -1.5946 | ||
| qERA05.2 | 18TS | A05 | 6.00 | me14em5-116—PM418 | 3.96 | 5.92 | -1.3923 | |
| qERA08.1 | 17BD | A08 | 7.69 | HBAUAh177—AhTE0658 | 3.01 | 3.30 | -0.8612 | |
| qERA08.2 | 18BD | A08 | 8.40 | HBAUAh177—AhTE0658 | 5.18 | 5.53 | -1.3478 |
表5
不同性状在相同标记区间的QTL分析结果"
| 染色体 Chr. | 标记区间 Marker interval | QTL | LOD | 贡献率 PVE (%) | |||
|---|---|---|---|---|---|---|---|
| A04 | GM1867-A04—AHGS1967-A04 | qMSHA04.1 | qMSHA04.2 | qMSHA04.3 | qMSHA04.4 | 3.11—7.49 | 3.25—10.05 |
| qMSHA04.5 | qMSHA04.6 | ||||||
| A05 | me14em5-116—PM418-A05 | qLBAA05.1 | qLBAA05.2 | qMSHA05.1 | qMSHA05.2 | 3.16—10.15 | 3.86—15.08 |
| qIOPTA05.1 | qIOPTA05.2 | qIOPTA05.3 | qERA05.1 | ||||
| qERA05.2 | |||||||
| A08 | HBAUAh177—AhTE0658-A08 | qLBLA08.1 | qLBLA08.2 | qERA08.1 | qERA08.2 | 3.01—6.13 | 3.30—8.56 |
表6
株型相关性状的上位性QTL定位结果"
| 性状 Trait | 上位性QTL Epi-QTL | 环境 Env. | 染色体 Chr. | 位置 Position | 标记区间 Marker interval | 染色体 Chr. | 位置 Position | 标记区间 Marker interval | LOD | 贡献率 PVE (%) | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 侧枝长 LBL | Epi-qLBLA04.1 | 18BD | A04 | 15.000 | ARS772—GM1867 | A04 | 20.000 | GM1867—AHGS1967 | 13.760 | 2.420 | |
| Epi-qLBLA02.1 | 18BD | A02 | 35.000 | AHGS1163—AHGS1886 | A02 | 40.000 | AHGS1886—AHGS1159 | 9.620 | 2.230 | ||
| Epi-qLBLA05.1 | 18BD | A05 | 35.000 | AhTE0588—T5me8-66 | A05 | 40.000 | T5me8-66—T2me4-75 | 10.420 | 2.250 | ||
| Epi-qLBLA03.1 | 18BD | A03 | 25.000 | GM1883—AhTE0570 | A03 | 30.000 | AhTE0570—TC4G02 | 11.900 | 2.280 | ||
| Epi-qLBLB03.1 | 18BD | B03 | 7.165 | AHBGSC1003E10—GM1996 | B03 | 8.609 | AHBGSC1003E10—GM1996 | 9.750 | 2.230 | ||
| Epi-qLBLA08.1 | 18BD | A08 | 6.255 | HBAUAh177—AhTE0658 | A08 | 10.000 | AhTE0658—TC6H03 | 7.810 | 2.780 | ||
| 株型 指数 IOPT | Epi-qIOPTB04.1 | 16BD | B04 | 64.814 | me2em16-141—me9em4-125 | B04 | 69.814 | me2em16-141—me9em4-125 | 8.920 | 1.360 | |
| Epi-qIOPTB07.1 | 16BD | B07 | 5.000 | TC1A08—TC9H09 | B07 | 10.000 | TC1A08—TC9H09 | 7.280 | 1.200 | ||
| Epi-qIOPTA02.4 | 16BD | A02 | 45.000 | AHGS1886—AHGS1159 | A02 | 50.000 | AHGS1886—AHGS1159 | 8.510 | 1.420 | ||
| Epi-qIOPTA03.4 | 16BD | A03 | 40.000 | AhTE0570—TC4G02 | A03 | 45.000 | AhTE0570—TC4G02 | 8.530 | 1.160 | ||
| Epi-qIOPTB03.1 | 16BD | B03 | 7.165 | AHBGSC1003E10—GM1996 | B03 | 17.351 | GM1996—seq2H08 | 8.760 | 1.440 | ||
| Epi-qIOPTA10.2 | 16BD | A10 | 20.000 | AhTE0586—AHGS1192 | A10 | 25.000 | AHGS1192—seq3e10 | 5.500 | 0.850 | ||
| Epi-qIOPTA04.5 | 17BD | A04 | 50.000 | GM1135—pPGSseq15C12a | A04 | 55.000 | GM1135—pPGSseq15C12a | 8.300 | 0.800 | ||
| Epi-qIOPTB07.3 | 17BD | B07 | 10.000 | TC1A08—TC9H09 | B07 | 15.000 | TC1A08—TC9H09 | 9.440 | 0.860 | ||
| Epi-qIOPTA02.3 | 17BD | A02 | 40.000 | AHGS1886—AHGS1159 | A02 | 45.000 | AHGS1886—AHGS1159 | 8.670 | 0.850 | ||
| Epi-qIOPTA08.1 | 17BD | A08 | 10.827 | me3em14-196—Ah4-4 | A08 | 15.544 | me3em14-196—Ah4-4 | 8.720 | 0.920 | ||
| Epi-qIOPTA05.1 | 17BD | A05 | 5.000 | me7em9-96—me13em5-112 | A05 | 10.000 | me7em9-96—me13em5-112 | 6.020 | 0.810 | ||
| Epi-qIOPTA03.1 | 17BD | A03 | 25.000 | GM1883—AhTE0570 | A03 | 35.000 | AhTE0570—TC4G02 | 7.840 | 0.720 | ||
| Epi-qIOPTA06.1 | 17BD | A06 | 10.164 | me4em15-95—me7em1-83 | A06 | 15.078 | me4em15-95—me7em1-83 | 7.270 | 0.830 | ||
| Epi-qIOPTB03.2 | 17BD | B03 | 7.165 | AHBGSC1003E10—GM1996 | B03 | 17.351 | GM1996—seq2H08 | 8.660 | 0.820 | ||
| Epi-qIOPTA10.3 | 17BD | A10 | 20.000 | AhTE0586—AHGS1192 | A10 | 25.000 | AHGS1192—seq3e10 | 7.930 | 0.780 | ||
| 性状 Trait | 上位性QTL Epi-QTL | 环境 Env. | 染色体 Chr. | 位置 Position | 标记区间 Marker interval | 染色体 Chr. | 位置 Position | 标记区间 Marker interval | LOD | 贡献率 PVE (%) | |
| Epi-qIOPTB07.6 | 17HD | B09 | 10.000 | T3me2-388—AHGS1576 | B09 | 15.000 | T3me2-388—AHGS1576 | 10.480 | 1.360 | ||
| Epi-qIOPTA04.4 | 17HD | A04 | 40.000 | AHGS1967—GM1135 | A04 | 50.000 | GM1135—pPGSseq15C12a | 11.580 | 1.280 | ||
| Epi-qIOPTB07.4 | 17HD | B07 | 10.000 | TC1A08—TC9H09 | B07 | 15.000 | TC1A08—TC9H09 | 10.810 | 1.330 | ||
| Epi-qIOPTA02.5 | 17HD | A02 | 50.000 | AHGS1886—AHGS1159 | A02 | 55.000 | AHGS1886—AHGS1159 | 10.100 | 1.410 | ||
| Epi-qIOPTA05.2 | 17HD | A05 | 5.000 | me14em5-116—PM418 | A05 | 10.000 | PM418—seq11E11 | 9.100 | 1.380 | ||
| Epi-qIOPTA08.4 | 17HD | A08 | 15.543 | me3em14-196—Ah4-4 | A08 | 20.260 | me3em14-196—Ah4-4 | 11.350 | 1.360 | ||
| Epi-qIOPTA05.3 | 17HD | A05 | 5.000 | me7em9-96—me13em5-112 | A05 | 10.000 | me7em9-96—me13em5-112 | 11.390 | 1.300 | ||
| Epi-qIOPTB07.5 | 17HD | B08 | 20.000 | me4em5-88—AI119H15 | B08 | 25.000 | me4em5-88—AI119H15 | 8.550 | 1.230 | ||
| Epi-qIOPTA03.2 | 17HD | A03 | 30.000 | AhTE0570—TC4G02 | A03 | 35.000 | AhTE0570—TC4G02 | 11.840 | 1.280 | ||
| Epi-qIOPTB03.3 | 17HD | B03 | 7.165 | AHBGSC1003E10—GM1996 | B03 | 8.609 | AHBGSC1003E10—GM1996 | 14.290 | 1.430 | ||
| Epi-qIOPTA08.2 | 17HD | A08 | 15.000 | AhTE0658—TC6H03 | A08 | 20.000 | TC6H03—AhTE0477 | 5.460 | 1.000 | ||
| Epi-qIOPTA04.2 | 17HD | A04 | 25.000 | GM1867—AHGS1967 | A08 | 25.000 | TC6H03—AhTE0477 | 5.640 | 0.480 | ||
| Epi-qIOPTA10.4 | 17HD | A10 | 20.000 | AhTE0586—AHGS1192 | A10 | 25.000 | AHGS1192—seq3e10 | 11.310 | 1.200 | ||
| Epi-qIOPTA04.3 | 18BD | A04 | 25.000 | GM1867—AHGS1967 | A04 | 30.000 | GM1867—AHGS1967 | 27.990 | 0.900 | ||
| Epi-qIOPTA02.1 | 18BD | A02 | 20.000 | Ah3TC13E05—AHGS1463 | A02 | 25.000 | AHGS1463—AHGS1163 | 25.850 | 0.760 | ||
| Epi-qIOPTA05.5 | 18BD | A05 | 25.000 | seq11E11—AhTE0588 | A05 | 30.000 | AhTE0588—T5me8-66 | 25.060 | 0.770 | ||
| Epi-qIOPTA03.5 | 18BD | A03 | 40.000 | AhTE0570—TC4G02 | A03 | 45.000 | AhTE0570—TC4G02 | 25.170 | 1.230 | ||
| Epi-qIOPTA06.2 | 18BD | A06 | 15.078 | me4em15-95—me7em1-83 | A06 | 20.000 | me7em1-83—me8em16-92 | 25.390 | 0.780 | ||
| Epi-qIOPTB03.5 | 18BD | B03 | 31.542 | seq2H08—GM1954 | B03 | 31.236 | GM1954—IPAHM103 | 24.970 | 0.770 | ||
| Epi-qIOPTA10.1 | 18BD | A10 | 15.000 | AhTE0586—AHGS1192 | A10 | 20.000 | AhTE0586—AHGS1192 | 21.990 | 0.770 | ||
| Epi-qIOPTB07.7 | 18HD | B09 | 15.000 | T3me2-388—AHGS1576 | B09 | 20.000 | T3me2-388—AHGS1576 | 13.540 | 0.870 | ||
| Epi-qIOPTA04.6 | 18HD | A04 | 70.000 | pPGSseq15C12a—me7em17-133 | A04 | 75.000 | pPGSseq15C12a—me7em17-133 | 18.600 | 0.850 | ||
| Epi-qIOPTB07.2 | 18HD | B07 | 5.000 | TC1A08—TC9H09 | B07 | 10.000 | TC1A08—TC9H09 | 13.000 | 0.890 | ||
| Epi-qIOPTA02.2 | 18HD | A02 | 35.000 | AHGS1163—AHGS1886 | A02 | 45.000 | AHGS1886—AHGS1159 | 14.530 | 0.850 | ||
| Epi-qIOPTA05.6 | 18HD | A05 | 50.000 | T2me4-75—seq18C2 | A05 | 55.000 | T2me4-75—seq18C2 | 16.160 | 0.950 | ||
| Epi-qIOPTA08.5 | 18HD | A08 | 20.260 | me3em14-196—Ah4-4 | A08 | 30.000 | Ah4-4—TC9B08 | 12.330 | 0.830 | ||
| Epi-qIOPTA05.4 | 18HD | A05 | 15.000 | me7em9-96—me13em5-112 | A05 | 20.000 | me7em9-96—me13em5-112 | 15.600 | 1.120 | ||
| Epi-qIOPTA03.6 | 18HD | A03 | 45.000 | AhTE0570—TC4G02 | A03 | 50.000 | TC4G02—IPAHM93 | 15.850 | 0.840 | ||
| Epi-qIOPTA06.3 | 18HD | A06 | 35.000 | me7em1-83—me8em16-92 | A06 | 40.000 | me8em16-92—AHGS1337 | 15.200 | 0.840 | ||
| Epi-qIOPTB03.4 | 18HD | B03 | 31.236 | GM1954—IPAHM103 | B03 | 45.139 | GM1954—IPAHM103 | 19.630 | 0.840 | ||
| Epi-qIOPTA08.3 | 18HD | A08 | 15.000 | AhTE0658—TC6H03 | A08 | 20.000 | TC6H03—AhTE0477 | 17.200 | 0.830 | ||
| Epi-qIOPTA10.5 | 18HD | A10 | 20.000 | AhTE0586—AHGS1192 | A10 | 25.000 | AHGS1192—seq3e10 | 19.290 | 0.830 | ||
| Epi-qIOPTA04.1 | 18TS | A04 | 5.000 | ARS772—GM1867 | A04 | 10.000 | ARS772—GM1867 | 5.060 | 0.260 | ||
| Epi-qIOPTA08.6 | 18TS | A08 | 20.260 | me3em14-196—Ah4-4 | A08 | 25.000 | Ah4-4—TC9B08 | 5.410 | 0.260 | ||
| Epi-qIOPTA03.3 | 18TS | A03 | 30.000 | AhTE0570—TC4G02 | A03 | 35.000 | AhTE0570—TC4G02 | 6.150 | 0.250 | ||
| 扩展 半径 ER | Epi-qERA09.1 | 17BD | A09 | 25.976 | AHTE0122—HBAUAh098 | A08 | 10.827 | me3em14-196—Ah4-4 | 5.830 | 12.110 | |
| Epi-qERB04.2 | 18HD | B04 | 64.814 | me2em16-141—me9em4-125 | B08 | 5.000 | AHGS1286—TC20B05 | 5.880 | 12.250 | ||
| Epi-qERB04.1 | 18TS | B04 | 24.814 | AHTE0445—PM36 | B04 | 29.814 | PM36—PMc348 | 6.590 | 7.280 | ||
| [1] | 张新友. 栽培花生产量、品质和抗病性的遗传分析与QTL定位研究[D]. 杭州: 浙江大学, 2011. |
| ZHANG X Y. Inheritance of main traits related to yield, quality and disease resistance and their QTLs mapping in peanut (Arachis hypogaea L.)[D]. Hangzhou: Zhejiang University, 2011. (in Chinese) | |
| [2] |
HAKE A A, KENTA S, ARATI Y, SUKRUTH M, MALAGOUDA P, NAYAK S N, LINGARAJU S, PATIL P V, NADAF H L, GOWDA M V C, BHAT R S. Mapping of important taxonomic and productivity traits using genic and non-genic transposable element markers in peanut (Arachis hypogaea L.). PLoS ONE, 2017,12(10):e0186113.
doi: 10.1371/journal.pone.0186113 pmid: 29040293 |
| [3] |
LI Y J, LI L Z, ZHANG X R, ZHANG K, MA D C, LIU J Q, WANG X J, LIU F Z. QTL mapping and marker analysis of main stem height and the first lateral branch length in peanut (Arachis hypogaea L.). Euphytica, 2017,213(2):57.
doi: 10.1007/s10681-017-1847-y |
| [4] |
LÜ J W, LIU N, GUO J B, XU Z J, LI X P, LI Z D, LUO H Y, REN X P, HUANG L, ZHOU X J, CHEN Y N, CHEN W G, LEI Y, TU J X, JIANG H F, LIAO B S. Stable QTLs for plant height on chromosome A09 identified from two mapping populations in peanut (Arachis hypogaea L.). Frontiers in Plant Science, 2018,9:684.
doi: 10.3389/fpls.2018.00684 pmid: 29887872 |
| [5] |
Wang Z H, Huai D X, Zhang Z H, Cheng K, Kang Y P, Wan L Y, Yan L Y, Jiang H F, Lei Y, Liao B S. Development of a high-density genetic map based on specific length amplified fragment sequencing, and its application in quantitative trait loci analysis for yield-related traits in cultivated peanut. Frontiers in Plant Science, 2018,9:827.
doi: 10.3389/fpls.2018.00827 pmid: 29997635 |
| [6] | 姜慧芳. 花生种质资源描述规范和数据标准3-9. 北京: 中国农业出版社, 2006. |
| JIANG H F. Specification for Description and Data of Peanut Germplasm Resources 3-9. Beijing: China Agriculture Press, 2006. (in Chinese) | |
| [7] | KAYAM G, BRAND Y, FAIGENBOIM D A, PATIL A, HEDVAT I, HOVAV R. Fine-mapping the branching habit trait in cultivated peanut by combining bulked segregant analysis and high-throughput sequencing. Frontiers in Plant Science, 2017,8(467):1-11. |
| [8] | 曹敏建, 王晓光, 于海秋. 花生:历史·栽培·育种·加工. 沈阳: 辽宁科学技术出版社, 2013. |
| CAO M J, WANG X G, YU H Q. Peanut: History・Growth・Breeding・Process. Shenyang: Liaoning Science and Techinology Publishing House, 2013. (in Chinese) | |
| [9] | 蓝新隆, 唐兆秀, 徐日荣. 福建花生产量与主要农艺性状之间的灰色关联度分析. 江西农业学报, 2011,23(8):61-63. |
| LAN X L, TANG Z X, XU R R. Analysis of gray correlation between yield and major agronomic traits of peanut in Fujian province. Acta Agriculturae Jiangxi, 2011,23(8):61-63. (in Chinese) | |
| [10] |
FONCEKA D, TOSSIM H A, RIVALLAN R, VIGNES H, LACUT E, BELLIS F, FAYE I, NDOYE O, SORAYA C M L B, JOSE ́F M V, DAVID J B, GLASZMANN J C, COURTOIS B, RAMI J F. Construction of chromosome segment substitution lines in peanut (Arachis hypogaea L.) using a wild synthetic and QTL mapping for plant morphology. PLoS ONE, 2012,7(11):e48642.
doi: 10.1371/journal.pone.0051130 pmid: 23226478 |
| [11] |
SHIRASAWA K, KOILKONDA P, AOKI K, HIRAKAWA H, TABATA S, WATANABE M, HASEGAWA M, KIYOSHIMA H, SUZUKI S, KUWATA C, NAITO Y, KUBOYAMA T, NAKAYA A, SASAMOTO S, WATANABE A, KATO M, KAWASHIMA K, KISHIDA Y, KOHARA M, KURABAYASHI A, CHIKA T, TSURUOKA H, WADA T, ISOBE S. In silico polymorphism analysis for the development of simple sequence repeat and transposon markers and construction of linkage map in cultivated peanut. BMC Plant Biology, 2012,12:80.
doi: 10.1186/1471-2229-12-80 pmid: 22672714 |
| [12] |
HUANG L, HE H, CHEN W G, REN X P, CHEN Y N, ZHOU X J, XIA Y L, WANG X L, JIANG X J, LIAO B S, JIANG H F. Quantitative trait locus analysis of agronomic and quality-related traits in cultivated peanut (Arachis hypogaea L.). Theoretical and Applied Genetics, 2015,128(6):1103-1115.
doi: 10.1007/s00122-015-2493-1 pmid: 25805315 |
| [13] |
LI L, YANG X L, CUI S L, MENG X H, MU G J, HOU M Y, HE M J, ZHANG H, LIU L F, CHEN C Y. Construction of high-density genetic map and mapping quantitative trait loci for growth habit-related traits of peanut (Arachis hypogaea L.). Frontiers in Plant Science, 2019,10:745.
doi: 10.3389/fpls.2019.00745 pmid: 31263472 |
| [14] | 王亮, 杨鑫雷, GETAHUN Addisu, 崔顺立, 穆国俊, 刘立峰, 李自超. 栽培种花生AFLP标记体系的优化及多态性引物筛选. 核农学报, 2017,31(11):2087-2095. |
| WANG L, YANG X L, GETAHUN A, CUI S L, MU G J, LIU L F, LI Z C. Screening for polymorphic primer pairs and optimization of AFLP marker system in peanut. Journal of Nuclear Agricultural Sciences, 2017,31(11):2087-2095. (in Chinese) | |
| [15] |
LIN Z X, HE D H, ZHANG X L, NIE Y C, GUO X P, FENG C D, STEWART J M. Linkage map construction and mapping QTL for cotton fibre quality using SRAP, SSR and RAPD. Plant Breeding, 2008,124(2):180-187.
doi: 10.1111/pbr.2005.124.issue-2 |
| [16] |
YU J W, YU S X, LU C R, WANG W, FAN S L, SONG M Z, LIN Z X, ZHANG X L, ZHANG J F. High-density linkage map of cultivated allotetraploid cotton based on SSR, TRAP, SRAP and AFLP markers. Journal of Integrative Plant Biology, 2007,49(5):716-724.
doi: 10.1111/j.1744-7909.2007.00459.x |
| [17] | 崔顺立, 刘立峰, 陈焕英, 耿立格, 孟成生, 杨余. 河北省花生地方品种基于SSR标记的遗传多样性. 中国农业科学, 2009,42(9):3346-3353. |
| CUI S L, LIU L F, CHEN H Y, GENG L G, MENG C S, YANG Y. Genetic diversity of peanut landraces in Hebei province revealed by SSR markers. Scientia Agricultura Sinica, 2009,42(9):3346-3353. (in Chinese) | |
| [18] | JoinMap 4.0 Software for the calculation of genetic linkage maps in experimental populations. Wageningen: Kyazma B. V; 2006. |
| [19] | KOSAMBI D D. The estimation of map distances from recombination values. Annals of Human Genetics, 2011; 1:172-175. |
| [20] | VOORRIPS R E. MapChart: Software for the graphical presentation of linkage maps and QTLs. Journal of Heredity, 2002; 1:77-78. |
| [21] |
MENG L, LI H H, ZHANG L Y, WANG J K. QTL IciMapping: Integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental populations. The Crop Journal, 2015,3(3):269-283.
doi: 10.1016/j.cj.2015.01.001 |
| [22] | 吕维娜. 花生栽培种SSR遗传连锁图谱构建及重要产量性状QTL定位分析[D]. 郑州: 郑州大学, 2014. |
| LÜ W N. Contruction of genetic linkage map based on SSR markers and QTLs identification for major yield traits in the cultivated peanut (Arachis hypogaea L.)[D]. Zhengzhou: Zhengzhou University, 2014. (in Chinese) | |
| [23] |
HAKE A A, SHIRASAWA K, YADAWAD A, SUKRUTH M, PATIL M, NAYAK S N. Mapping of important taxonomic and productivity traits using genic and non-genic transposable element markers in peanut (Arachis hypogaea L.). PLoS ONE, 2017,12:e0186113.
doi: 10.1371/journal.pone.0186113 pmid: 29040293 |
| [24] |
LI Y, LI L, ZHANG X, ZHANG K, MA D, LIU J. QTL mapping and marker analysis of main stem height and the first lateral branch length in peanut (Arachis hypogaea L.). Euphytica, 2017,213:57.
doi: 10.1007/s10681-017-1847-y |
| [25] |
DONALD C M. The breeding of crop ideotypes. Euphytica, 1968,17:385-403.
doi: 10.1007/BF00056241 |
| [26] |
成良强, 唐梅, 任小平, 黄莉, 陈伟刚, 李振动, 周小静, 陈玉宁, 廖伯寿, 姜慧芳. 栽培种花生遗传图谱的构建及主茎高和总分枝数QTL分析. 作物学报, 2015,41(6):979-987.
doi: 10.3724/SP.J.1006.2015.00979 |
|
CHENG L Q, TANG M, REN X P, HUANG L, CHEN W G, LI Z D, ZHOU X J, CHEN Y N, LIAO B S, JIANG H F. Construction of genetic map and QTL analysis for mainstem height and total branch number in peanut (Arachis hypogaea L.). Acta Agronomica Sinica, 2015,41(6):979-987. (in Chinese)
doi: 10.3724/SP.J.1006.2015.00979 |
|
| [27] |
HUANG L, REN X P, WU B, LI X P, CHEN W G, ZHOU X J, CHEN Y N, PANDEY M K, JIAO Y Q, LUO H Y, LEI Y, VARSHNEY R K, GUO B Z, JIANG H F. Development and deployment of a high- density linkage map identified quantitative trait loci for plant height in peanut (Arachis hypogaea L.). Scientific Reports, 2016,6:39478.
doi: 10.1038/srep39478 pmid: 27995991 |
| [28] |
ZHOU X J, XIA Y L, LIAO J H, LIU K D, LI Q, DONG Y, REN X P, CHEN Y N, HUANG L, LIAO B S, LEI Y, YAN L Y, JIANG H F. Quantitative trait locus analysis of late leaf spot resistance and plant-type-related traits in cultivated peanut (Arachis hypogaea L.) under multi-environments. PLoS ONE, 2016,11(11):e0166873.
doi: 10.1371/journal.pone.0166873 pmid: 27870916 |
| [29] |
UPADHYAYA H D, NIGAM S N. Detection of epistasis for protein and oil contents and oil quality parameters in peanut. Crop Science, 1999,39(1):115-118.
doi: 10.2135/cropsci1999.0011183X003900010018x |
| [30] |
UPADHYAYA H D, NIGAM S N. Epistasis for vegetative and reproductive traits in peanut. Crop Science, 1998,38(1):44-49.
doi: 10.2135/cropsci1998.0011183X003800010008x |
| [1] | 陈吉浩, 周界光, 曲翔汝, 王素容, 唐华苹, 蒋云, 唐力为, $\boxed{\hbox{兰秀锦}}$, 魏育明, 周景忠, 马建. 四倍体小麦胚大小性状QTL定位与分析[J]. 中国农业科学, 2023, 56(2): 203-216. |
| [2] | 王彩香,袁文敏,刘娟娟,谢晓宇,马麒,巨吉生,陈炟,王宁,冯克云,宿俊吉. 西北内陆早熟陆地棉品种的综合评价及育种演化[J]. 中国农业科学, 2023, 56(1): 1-16. |
| [3] | 唐华苹,陈黄鑫,李聪,苟璐璐,谭翠,牟杨,唐力为,兰秀锦,魏育明,马建. 基于55K SNP芯片的普通小麦穗长非条件和条件QTL分析[J]. 中国农业科学, 2022, 55(8): 1492-1502. |
| [4] | 吴月,隋新华,戴良香,郑永美,张智猛,田云云,于天一,孙学武,孙棋棋,马登超,吴正锋. 慢生根瘤菌及其与花生共生机制研究进展[J]. 中国农业科学, 2022, 55(8): 1518-1528. |
| [5] | 赵凌, 张勇, 魏晓东, 梁文化, 赵春芳, 周丽慧, 姚姝, 王才林, 张亚东. 利用高密度Bin图谱定位水稻抽穗期剑叶叶绿素含量QTL[J]. 中国农业科学, 2022, 55(5): 825-836. |
| [6] | 王慧玲, 闫爱玲, 孙磊, 张国军, 王晓玥, 任建成, 徐海英. 鲜食葡萄果实单萜合成关键基因的eQTL分析[J]. 中国农业科学, 2022, 55(5): 977-990. |
| [7] | 卞能飞, 孙东雷, 巩佳莉, 王幸, 邢兴华, 金夏红, 王晓军. 花生烘烤食用品质评价及指标筛选[J]. 中国农业科学, 2022, 55(4): 641-652. |
| [8] | 刘进,胡佳晓,马小定,陈武,勒思,Jo Sumin,崔迪,周慧颖,张立娜,Shin Dongjin,黎毛毛,韩龙植,余丽琴. 水稻RIL群体高密度遗传图谱的构建及苗期耐热性QTL定位[J]. 中国农业科学, 2022, 55(22): 4327-4341. |
| [9] | 王娟,陈皓宁,石大川,于天一,闫彩霞,孙全喜,苑翠玲,赵小波,牟艺菲,王奇,李春娟,单世华. 花生高亲和硝酸盐转运蛋白基因AhNRT2.7a响应低氮胁迫的功能研究[J]. 中国农业科学, 2022, 55(22): 4356-4372. |
| [10] | 谢晓宇, 王凯鸿, 秦晓晓, 王彩香, 史春辉, 宁新柱, 杨永林, 秦江鸿, 李朝周, 马麒, 宿俊吉. 陆地棉吐絮率的限制性两阶段多位点全基因组关联分析及候选基因预测[J]. 中国农业科学, 2022, 55(2): 248-264. |
| [11] | 邹林翰,周新颖,张泽源,蔚睿,袁梦,宋晓朋,简俊涛,张传量,韩德俊,宋全昊. 小麦周8425B×小偃81重组自交系群体千粒重相关性状的QTL定位及单倍型分析[J]. 中国农业科学, 2022, 55(18): 3473-3483. |
| [12] | 郭灿,岳晓凤,白艺珍,张良晓,张奇,李培武. 花生黄曲霉毒素平衡取样-随机森林风险预警模型的应用研究[J]. 中国农业科学, 2022, 55(17): 3426-3436. |
| [13] | 常立国,何坤辉,刘建超. 多环境下玉米保绿相关性状遗传位点的挖掘[J]. 中国农业科学, 2022, 55(16): 3071-3081. |
| [14] | 郭淑青,宋慧,柴少华,郭岩,石兴,杜丽红,邢璐,解慧芳,张扬,李龙,冯佰利,刘金荣,杨璞. 谷子生育期及穗相关性状的QTL定位[J]. 中国农业科学, 2022, 55(15): 2883-2898. |
| [15] | 史晓龙,郭佩,任婧瑶,张鹤,董奇琦,赵新华,周宇飞,张正,万书波,于海秋. 基于花生//高粱间作模式的花生盐胁迫耐受性效应研究[J]. 中国农业科学, 2022, 55(15): 2927-2937. |
|
||