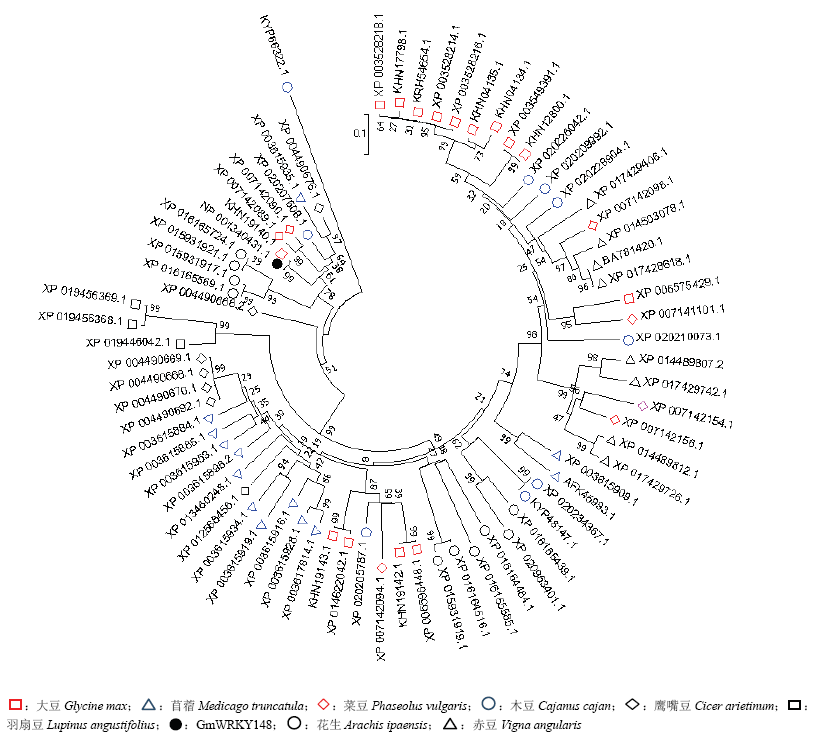
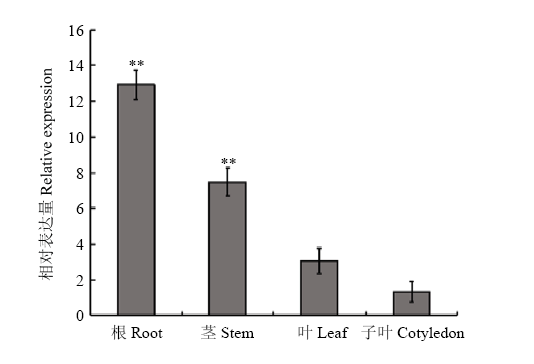
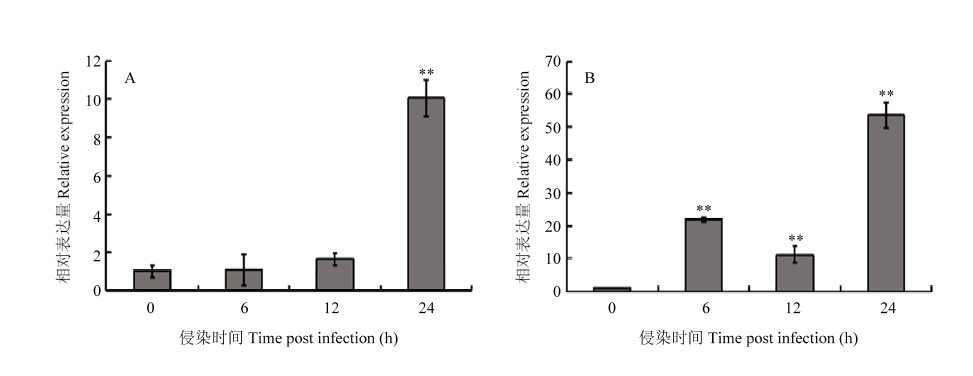
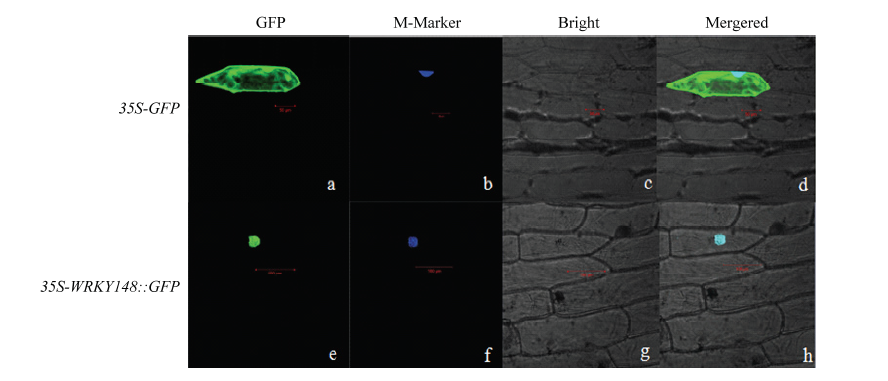
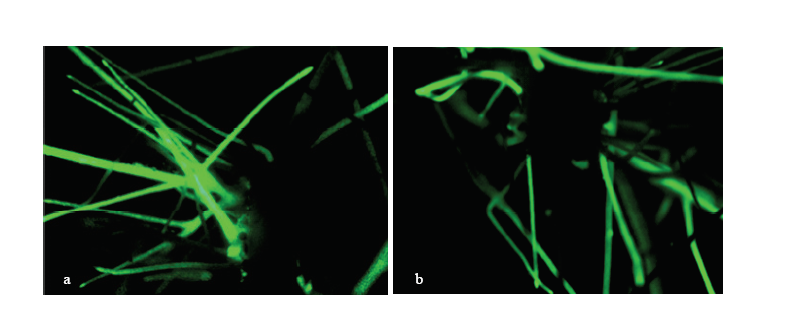
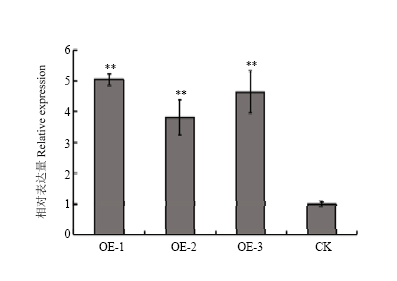
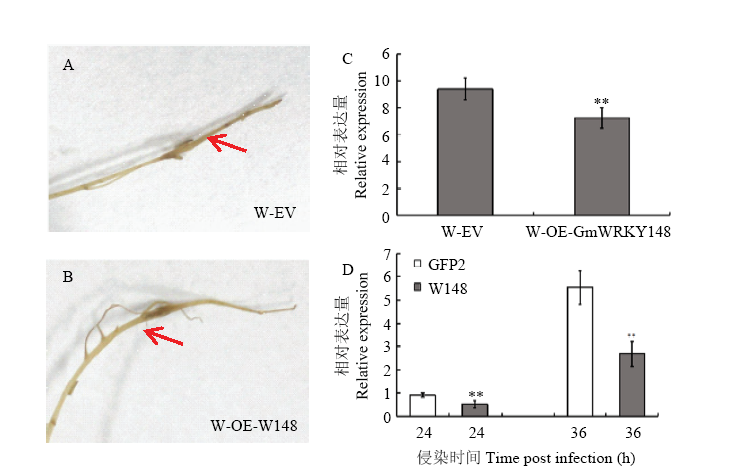
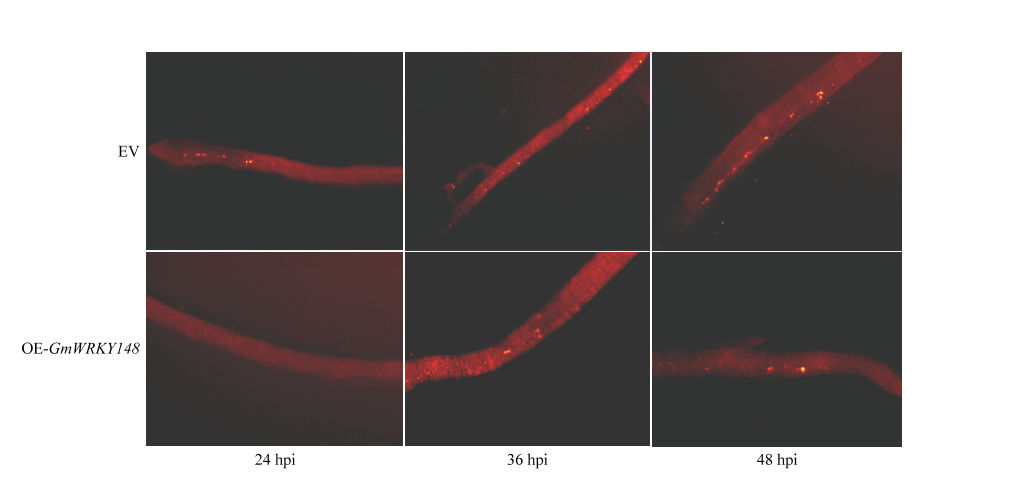
Scientia Agricultura Sinica ›› 2018, Vol. 51 ›› Issue (18): 3445-3454.doi: 10.3864/j.issn.0578-1752.2018.18.002
• CROP GENETICS & BREEDING·GERMPLASM RESOURCES·MOLECULAR GENETICS • Previous Articles Next Articles
ShaSha WANG( ), XiaoXia CUI, YanZhong HUANG, HuiDong XUAN, Na GUO, Han XING()
), XiaoXia CUI, YanZhong HUANG, HuiDong XUAN, Na GUO, Han XING()
| [1] | 张宝强. 大豆对大豆疫霉根腐病的抗源筛选及抗性遗传分析[D]. 南京: 南京农业大学, 2010. |
| ZHANG B B Q.Studies on screening for resistance source and genetic analysis to P. sojae in soybean [D]. Nanjing: Nanjing Agricultural University, 2010. (Chinese) | |
| [2] |
LI D, LIU P, YU J, DOSSA K, ZHANG Y, ZHOU R, WEI X, ZHANG X.Genome-wide analysis of WRKY gene family in the sesame genome and identification of the WRKY genes involved in responses to abiotic stresses.BMC Plant Biology, 2017, 17(1): 152.
doi: 10.1186/s12870-017-1099-y pmid: 5594535 |
| [3] |
RUSHTON P J, TORRES J T, PARNISKE M, WERNERT P, HAHIBROCK K, SOMSSICH I E.Interaction of elicitor-induced DNA-binding proteins with elicitor response elements in the promoters of parsley PR1 genes.The EMBO Journal, 1996, 15(20): 5690-5700.
doi: 10.1002/j.1460-2075.1996.tb00953.x pmid: 8896462 |
| [4] |
ROCHER A, DUMAS C, COCK J M.A W-box is required for full expression of the SA-responsive gene SFR2.Gene, 2005, 344(1): 181-192.
doi: 10.1016/j.gene.2004.09.016 pmid: 15656984 |
| [5] |
KIM C Y, ZHANG S.Activation of a mitogen-activated protein kinase cascade induces WRKY family of transcription factors and defense genes in tobacco.The Plant Journal, 2004, 38(1): 142-151.
doi: 10.1111/j.1365-313X.2004.02033.x pmid: 202020202020042020203820142151 |
| [6] |
PANDEY S P, SOMSSICH I E.The role of WRKY transcription factors in plant immunity.Plant Physiology, 2009, 150(4): 1648-1655.
doi: 10.1104/pp.109.138990 pmid: 19420325 |
| [7] |
BANERJEE A, ROYCHOUDHURY A.WRKY proteins: Signaling and regulation of expression during abiotic stress responses.The Scientific World Journal, 2015, 2015(807560): 807560.
doi: 10.1155/2015/807560 pmid: 4387944 |
| [8] |
HAN Y, ZHANG X, WANG Y, MING F.Correction: The suppression of WRKY44 by GIGANTEA- miR172 pathway is involved in drought response ofArabidopsis thaliana. PLOS ONE, 2015, 10(4): e0124854.
doi: 10.1371/journal.pone.0124854 pmid: 25844872 |
| [9] |
ZELICOURT A D, COLCOMBET J, HIRT H.The role of MAPK modules and ABA during abiotic stress signaling.Trends in Plant Science, 2016, 21(8): 677-685.
doi: 10.1016/j.tplants.2016.04.004 pmid: 27143288 |
| [10] | ASAI T, TENA G, PLOTNIKOVA J, WILLMANN M R, CHIU W L, LOURDES G G, BOLLER T, AUSUBEL F M, SHEEN J.MAP kinase signalling cascade inArabidopsis innate immunity. Nature, 2002, 415(6875): 977-983. |
| [11] | LI Y, WILLIAMS B, DICKMAN M.Arabidopsis B-cell lymphoma 2 (Bcl-2)-associated athano gene 7(BAG7)-mediated heat tolerance requires translocation, sumoylation and binding to WRKY29. New Phytologist, 2016, 214(2): 695-705. |
| [12] |
DANQUAH A, DE A Z, COLCOMBET J, HIRT H.The role of ABA and MAPK signaling pathways in plant abiotic stress responses.Biotechnology Advances, 2014, 32(1): 40.
doi: 10.1016/j.biotechadv.2013.09.006 |
| [13] | JIANG Y, LIANG G, YANG S, YU D.Arabidopsis WRKY57 functions as a node of convergence for jasmonic-acid and auxin- mediated signaling in jasmonic acid-induced leaf senescence. The Plant Cell, 2014, 26(1): 230-245. |
| [14] |
ZHANG Y, YU H, YANG X, LI Q, LING J, WANG H, GU X, HUANG S, JIANG W.CsWRKY46, a WRKY transcription factor from cucumber, confers cold resistance in transgenic-plant by regulating a set of cold-stress responsive genes in an ABA-dependent manner.Plant Physiology & Biochemistry, 2016, 108: 478.
doi: 10.1016/j.plaphy.2016.08.013 pmid: 27592172 |
| [15] |
JOSHI R, WANI S H, SINGH B, BOHRA A, DAR Z A, LONE A A, PAREE K A, SINGLA-PAREEK S L. Transcription factors and plants response to drought stress: Current understanding and future directions.Frontiers in Plant Science, 2016, 7(e102529): 1029.
doi: 10.3389/fpls.2016.01029 pmid: 27471513 |
| [16] |
OHAMA N, SATO H, SHINOZAKI K, YAMAGUCHI-SHINOZAKI K.Transcriptional regulatory network of plant heat stress response.Trends in Plant Science, 2017, 22(1): 53-65.
doi: 10.1016/j.tplants.2016.08.015 pmid: 27666516 |
| [17] |
GU L, LI L, WEI H, WANG H, SU J, GUO Y, YU S.Identification of the group IIa WRKY subfamily and the functional analysis of GhWRKY17 in upland cotton (Gossypium hirsutum L.). PLOS ONE, 2018, 13(1): e0191681.
doi: 10.1371/journal.pone.0191681 pmid: 5784973 |
| [18] |
WU X, SHIROTO Y, KISHITANI S, ITO Y, TORIYAMA K.Enhanced heat and drought tolerance in transgenic rice seedlings overexpressing OsWRKY11 under the control of HSP101 promoter.Plant Cell Reports, 2009, 28(1): 21-30.
doi: 10.1007/s00299-008-0614-x pmid: 2018818929 |
| [19] |
HE G H, XU J Y, WANG Y X, LIU J M, LI P S, CHEN M, MA Y Z, XU Z S.Drought-responsive WRKY transcription factor genes TaWRKY1, and TaWRKY33, from wheat confer drought and/or heat resistance inArabidopsis. BMC Plant Biology, 2016, 16(1): 116.
doi: 10.1186/s12870-016-0806-4 pmid: 27215938 |
| [20] | JIA H, WANG C, WANG F, LIU S, LI G, GUO X.GhWRKY68 reduces resistance to salt and drought in transgenic Nicotiana benthamiana. PLOS ONE, 2015, 10(3): e0120646. |
| [21] |
CHANG C, YU D, JIAO J, JING S, SCHULZE-LEFERT P, SHEN Q H.Barley MLA immune receptors directly interfere with antagonistically acting transcription factors to initiate disease resistance signaling.The Plant Cell, 2013, 25(3): 1158-1173.
doi: 10.1105/tpc.113.109942 pmid: 23532068 |
| [22] | DESLANDES L, OLIVIER J, THEULIERES F, HIRSCH J, FENG D X, BITTNER-EDDY P, BEYON J, MARCO Y.Resistance toRalstonia solanacearum in Arabidopsis thaliana is conferred by the recessive RRS1-R gene, a member of a novel family of resistance genes. Proceedings of the National Academy of Sciences of the United States of America, 2002, 99(4): 2404-2409. |
| [23] | 成洪涛, 王石平. 水稻-病原互作中的重要角色: WRKY类转录因子. 中国科学: 生命科学, 2014(8): 784-793. |
| CHENG H T, WANG S P.WRKY- type transcription factors: A significant factor in rice- pathogen interactions.Scientia Sinica, 2014, 44(8): 784-793.(in Chinese) | |
| [24] | WANG J, TAO F, AN F, ZOU Y P, TIAN W, CHEN X M, XU X M, HU X P.Wheat transcription factor TaWRKY70 is positively involved in high-temperature seedling-plant resistance toPuccinia striiformis f. sp. tritici. Molecular Plant Pathology, 2017, 18(5): 649-661. |
| [25] |
SUGIMOTO T, WATANABE K, YOSHIDA S, BIGGS A R.Field application of calcium to reduce Phytophthora stem rot of soybean, and calcium distribution in plants.Plant Disease, 2010, 94(7): 812-819.
doi: 10.1094/PDIS-94-7-0812 |
| [26] | 杨晓贺, 张瑜, 顾鑫, 赵海红, 姚亮亮, 刘伟, 申宏波, 丁俊杰. 大豆疫霉根腐病的综合防治. 大豆科学, 2014, 33(4): 554-558. |
| YANG X H, ZHANG Y, GU X, ZHAO H H, YAO L L, LIU W, SHEN H B, DING J J.Integrated control of soybean phytophthora root rot.Soybean Science, 2014, 33(4): 554-558. (in Chinese) | |
| [27] | 陶恺. 大豆疫霉侵染过程的分子细胞学研究[D]. 南京: 南京农业大学, 2012. |
| TAO K.Molecular and cellular biology of interactions between Phytophthora sojae and host soybean[D]. Nanjing: Nanjing Agricultural University, 2012. (in Chinese) | |
| [28] | MOY P, QUTOB D, CHAPMAN B P, ATKINSON I, GIJZEN M.Patterns of gene expression upon infection of soybean plants byPhytophthora sojae. Molecular Plant Microbe Interactions, 2004, 17(10): 1051-1062. |
| [29] |
DUNIWAY J M.Water relations of water molds. Annual Review of Phytopathology, 1979, 17(17): 431-460.
doi: 10.1146/annurev.py.17.090179.002243 |
| [30] |
NIKLAUS J G, WILBERT G F.The biology of phytophthora infestans at its center of origin.Annual Review of Phytopathology, 2005, 43(43): 171.
doi: 10.1146/annurev.phyto.43.040204.135906 |
| [31] | SINCLAIR J B.Compendium of Soybean Diseases. Urbana- Champaign:American Phytopathological Society and University of Illinois, 1982. |
| [32] |
HARDHAM A, GUBLER F.Polarity of attachment of zoospores of a root pathogen and pre- alignment of the emerging germ tube.Cell Biology International Reports, 1990, 14(11): 947-956.
doi: 10.1016/0309-1651(90)90107-A |
| [33] |
HARDHAM A R.The cell biology behind Phytophthora pathogenicity. Australasian Plant Pathology, 2001, 30(2): 91-98.
doi: 10.1071/AP01006 |
| [34] | 张鑫. 大豆疫霉G蛋白α亚基调控游动孢子趋化性的机制研究[D]. 南京: 南京农业大学, 2015. |
| ZHANG X.Molecular mechanisms of Phytophthora sojae PsGPA1 in regulation of zoospore chemotaxis to soybean[D]. Nanjing: Nanjing Agricultural University, 2015. (in Chinese) |
| [1] | YAN Qiang,XUE Dong,HU YaQun,ZHOU YanYan,WEI YaWen,YUAN XingXing,CHEN Xin. Identification of the Root-Specific Soybean GmPR1-9 Promoter and Application in Phytophthora Root-Rot Resistance [J]. Scientia Agricultura Sinica, 2022, 55(20): 3885-3896. |
| [2] | ZaiDong LIU,Shan MENG,JianBo HE,GuangNan XING,WuBin WANG,TuanJie ZHAO,JunYi GAI. A Comparative Study on Linkage and Association QTL Mapping for Seed Isoflavone Contents in a Recombinant Inbred Line Population of Soybean [J]. Scientia Agricultura Sinica, 2020, 53(9): 1756-1772. |
| [3] | ShuGuang LI,YongCe CAO,JianBo HE,WuBin WANG,GuangNan XING,JiaYin YANG,TuanJie ZHAO,JunYi GAI. Genetic Dissection of Protein Content in a Nested Association Mapping Population of Soybean [J]. Scientia Agricultura Sinica, 2020, 53(9): 1743-1755. |
| [4] | LiYuan PAN,JianBo HE,JinMing ZHAO,WuBin WANG,GuangNan XING,DeYue YU,XiaoYan ZHANG,ChunYan LI,ShouYi CHEN,JunYi GAI. Detection Power of RTM-GWAS Applied to 100-Seed Weight QTL Identification in a Recombinant Inbred Lines Population of Soybean [J]. Scientia Agricultura Sinica, 2020, 53(9): 1730-1742. |
| [5] | XiaoShuai HAO,MengMeng FU,ZaiDong LIU,JianBo HE,YanPing WANG,HaiXiang REN,DeLiang WANG,XingYong YANG,YanXi CHENG,WeiGuang DU,JunYi GAI. Genome-Wide QTL-Allele Dissection of 100-Seed Weight in the Northeast China Soybean Germplasm Population [J]. Scientia Agricultura Sinica, 2020, 53(9): 1717-1729. |
| [6] | HAO QingTing,ZHANG Fei,JI XiaJie,XUE JinAi,LI RunZhi. Phenotypic Analysis of Epoxygenase-Transgenic Soybeans [J]. Scientia Agricultura Sinica, 2019, 52(2): 191-200. |
|
||