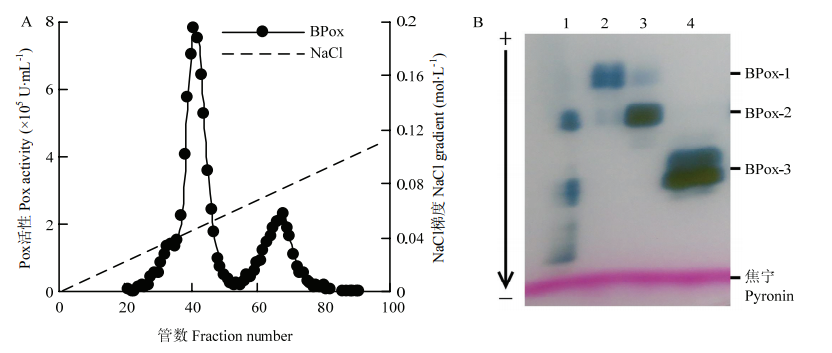
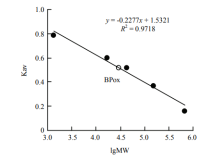
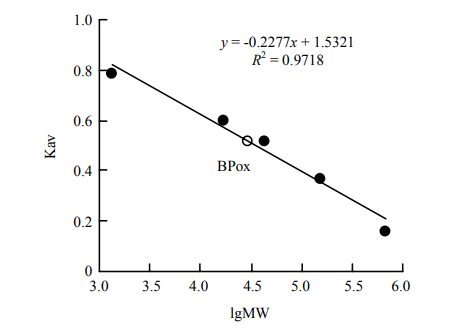
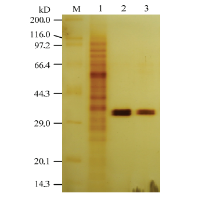
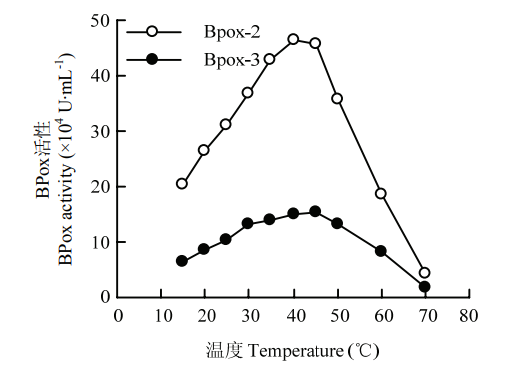
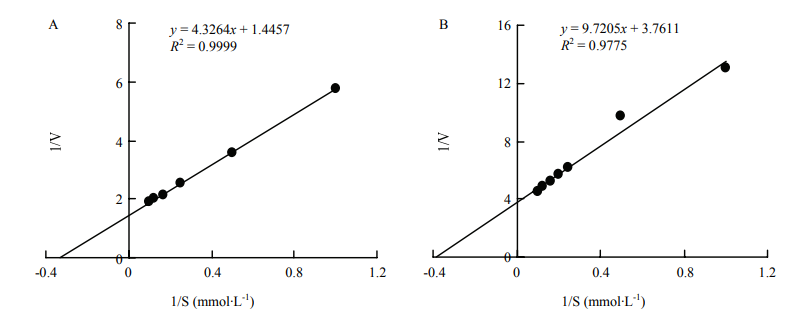
| [1] |
HIRAGA S, SASAKI K, ITO H, OHASHI Y, MATSUI H. A large family of class III plant peroxidases. Plant and Cell Physiology, 2001, 42(5):462-468.
doi: 10.1093/pcp/pce061
|
| [2] |
COSIO C, DUNAND C. Specific functions of individual class III peroxidase genes. Journal of Experimental Botany, 2008, 60(2):391-408.
doi: 10.1093/jxb/ern318
|
| [3] |
ZIPOR G, OREN-SHAMIR M. Do vacuolar peroxidases act as plant caretakers? Plant Science, 2013, 199/200:41-47.
doi: 10.1016/j.plantsci.2012.09.018
|
| [4] |
RING L, YEH S Y, HÜCHERIG S, HOFFMANN T, BLANCO- PORTALES R, FOUCHE M, VILLATORO C, DENOYES B, MONFORT A, CABALLERO J L, MUÑOZ-BLANCO J, GERSHENSON J, SCHWAB W. Metabolic interaction between anthocyanin and lignin biosynthesis is associated with peroxidase FaPRX27 in strawberry fruit. Plant Physiology, 2013, 163(1):43-60.
doi: 10.1104/pp.113.222778
|
| [5] |
YEH S Y, HUANG F C, HOFFMANN T, MAYERSHOFER M, SCHWAB W. FaPOD27 functions in the metabolism of polyphenols in strawberry fruit (Fragaria sp.). Frontiers in Plant Science, 2014, 5:518.
|
| [6] |
田国忠, 李怀方, 裘维蕃. 植物过氧化物酶研究进展. 武汉植物学研究, 2001, 19(4):332-344.
|
|
TIAN G Z, LI H F, QIU W F. Advances on research of plant peroxidases. Journal of Wuhan Botanical Research, 2001, 19(4):332-344. (in Chinese)
|
| [7] |
UNDERHILL S J R, CRITCHLEY C. Cellular localisation of polyphenol oxidase and peroxidase activity in Litchi chinensis Sonn. Pericarp. Australian Journal of Plant Physiology, 1995, 22(4):627-632.
|
| [8] |
GONG Q Q, TIAN S P. Partial characterization of soluble peroxidase in pericarp of litchi fruit. Progress in Biochemistry and Biophysics, 2002, 29(6):891-896.
|
| [9] |
JIANG Y, DUAN X, JOYCE D, ZHANG Z, Li J. Advances in understanding of enzymatic browning in harvested litchi fruit. Food Chemistry, 2004, 88:443-446.
doi: 10.1016/j.foodchem.2004.02.004
|
| [10] |
庞学群, 段学武, 张昭其, 徐凤彩, 季作梁. 荔枝果皮过氧化物酶的纯化及部分酶学性质研究. 热带亚热带植物学报, 2004, 12(5):449-454.
|
|
PANG X Q, DUAN X W, ZHANG Z Q, XU F C, JI Z L. Purification and some properties of peroxidase from pericarp of litchi (Litchi chinensis Sonn.). Journal of Tropical and Subtropical Botany, 2004, 12(5):449-454. (in Chinese)
|
| [11] |
ZHANG Z Q, PANG X Q, DUAN X W, JI Z L, JIANG Y M. Role of peroxidase in anthocyanin degradation in litchi fruit pericarp. Food Chemistry, 2005, 90:47-52.
doi: 10.1016/j.foodchem.2004.03.023
|
| [12] |
郑雯, 张永丽, 王家保, 金志强. 荔枝果皮2个POD同源基因的生物信息学分析及酶活测定. 热带作物学报, 2011, 32(3):437-442.
|
|
ZHENG W, ZHANG Y L, WANG J B, JIN Z Q. Bioinformatics analysis and function forecast of two POD genes in Litchi chinensis pericarp. Chinese Journal of Tropical Crops, 2011, 32(3):437-442. (in Chinese)
|
| [13] |
REICHEL M, TRIANI R, WELLHÖFER J, SRUAMSIRI P, CARLE R, NEIDHART S. Vital characteristics of litchi (Litchi chinensis Sonn.) pericarp that define postharvest concepts for Thai cultivars. Food and Bioprocess Technology, 2013, 6(5):1191-1206.
doi: 10.1007/s11947-011-0762-9
|
| [14] |
吴振先, 苏美霞, 陈维信, 胡桂兵. 贮藏荔枝果皮多酚氧化酶及过氧化物酶与褐变的研究. 华南农业大学学报, 1998, 19(1):12-15.
|
|
WU Z X, SU M X, CHEN W X, HU G B. Studies on polyphenol oxidase and peroxidase and litchi pericarp browning during storage. Journal of South China Agricultural University, 1998, 19(1):12-15. (in Chinese)
|
| [15] |
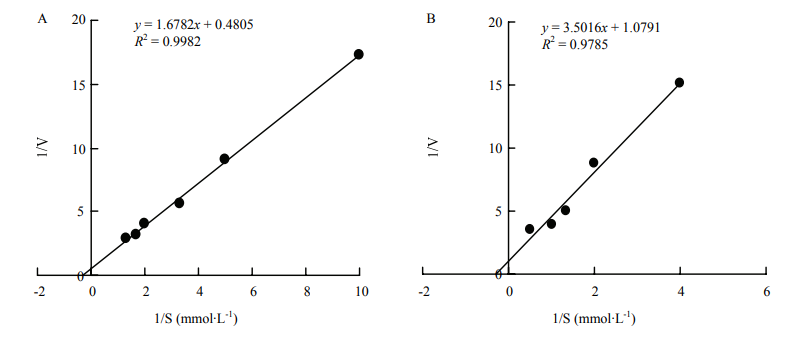
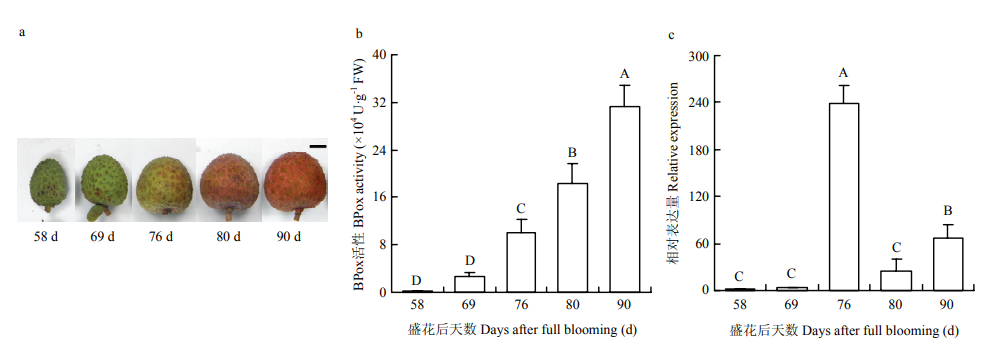
王蓓, 郭志雄, 柯思敏, 潘腾飞, 潘东明. 荔枝果皮的结合态POD及其在果实生长发育过程中的变化. 果树学报, 2014, 31(4):642-647, 754.
|
|
WANG B, GUO Z X, KE S M, PAN T F, PAN D M. Membrane/ wall-bound peroxidases in litchi pericarp and their changes during fruit development. Journal of Fruit Science, 2014, 31(4):642-647, 754.(in Chinese)
|
| [16] |
GUO Z X, PAN T F, LI K T, ZHONG F L, LIN L, PAN D M, LU L X. Cloning of NAD-SDH cDNA from plum fruit and its expression and characterization. Plant Physiology and Biochemistry, 2012, 57:175-180.
doi: 10.1016/j.plaphy.2012.05.021
|
| [17] |
BLUM H, BEIER H, GROSS H J. Improved silver staining of plant proteins, RNA and DNA in polyacrylamide gels. Electrophoresis, 1987, 8(2):93-99.
doi: 10.1002/(ISSN)1522-2683
|
| [18] |
李开拓. 荔枝果实成熟过程中的差异蛋白质组学研究[D]. 福州: 福建农林大学, 2011.
|
|
LI K T. Studies on differential proteomics during fruit ripening in litchi (Litchi chinensis Sonn.)[D]. Fuzhou: Fujian Agriculture and Forestry University, 2011. (in Chinese)
|
| [19] |
王蓓. 荔枝果皮可溶性POD组分的纯化、鉴定及其生化特性研究[D]. 福州: 福建农林大学, 2014.
|
|
WANG B. Purification, identification and characterization of soluble peroxidase isoforms in litchi pericarp[D]. Fuzhou: Fujian Agriculture and Forestry University, 2014. (in Chinese)
|
| [20] |
LIVAK K J, SCHMITTGEN T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCt Method. Methods, 2001, 25(4):402-408.
doi: 10.1006/meth.2001.1262
|
| [21] |
柯思敏. 荔枝结合态过氧化物酶分离纯化及其性质的初步研究[D]. 福州: 福建农林大学, 2015.
|
|
KE S M. A preliminary study on the purification and characterization of bound peroxidases in litchi pericarp[D]. Fuzhou: Fujian Agriculture and Forestry University, 2015. (in Chinese)
|
| [22] |
李开拓, 郭志雄, 潘东明, 钟凤林, 潘腾飞. 荔枝果皮总蛋白质提取及双向电泳体系的建立. 热带亚热带植物学报, 2011, 19(1):69-74.
|
|
LI K T, GUO Z X, PAN D M, ZHONG F L, PAN T F. Extraction of total protein from litchi pericarp and establishment of two-dimensional electrophoresis. Journal of Tropical and Subtropical Botany, 2011, 19(1):69-74. (in Chinese)
|
| [23] |
LAI B, HU B, QIN Y H, ZHAO J T, WANG H C, HU G B. Transcriptomic analysis of Litchi chinensis pericarp during maturation with a focus on chlorophyll degradation and flavonoid biosynthesis. BMC Genomics, 2015, 16:225.
doi: 10.1186/s12864-015-1433-4
|
| [24] |
ONSA G H, SAARI N B, SELAMAT J, BAKAR J. Purification and characterization of membrane-bound peroxidases from Metroxylon sagu. Food Chemistry, 2004, 85:365-376.
doi: 10.1016/j.foodchem.2003.07.013
|
| [25] |
SUN J, JIANG Y M, WEI X Y, SHI J, YOU Y L, LIU H, KAKUDA Y, ZHAO M M. Identification of (-)-epicatechin as the direct substrate for polyphenol oxidase isolated from litchi pericarp. Food Research International, 2006, 39:864-870.
doi: 10.1016/j.foodres.2006.05.001
|
| [26] |
LIU L, CAO S Q, XIE B J, SUN Z D, LI X Y, MIAO W H. Characterization of polyphenol oxidase from litchi pericarp using (-)-epicatechin as substrate. Journal of Agriculture and Food Chemistry, 2007, 55:7140-7143.
doi: 10.1021/jf070964a
|
| [27] |
MATSUI T, TABAYASHI A, IWANO M, SHINMYO A, KATO K, NAKAYAMA H. Activity of the C-terminal-dependent vacuolar sorting signal of horseradish peroxidase C1a is enhanced by its secondary structure. Plant and Cell Physiology, 2011, 52(2):413-420.
doi: 10.1093/pcp/pcq205
|
| [28] |
MATSUI T, NAKAYAMA H, YOSHIDA K, SHINMYO A. Vesicular transport route of horseradish C1a peroxidase is regulated by N- and C-terminal propeptides in tobacco cells. Applied Microbiology and Biotechnology, 2003, 62(5):517-522.
doi: 10.1007/s00253-003-1273-z
|
| [29] |
SCHNELL J A, HAN S Y, MIKI B L, JOHNSON D A. Soybean peroxidase propeptides are functional signal peptides and increase the yield of a foreign protein. Plant Cell Reports, 2010, 29(9):987-996.
doi: 10.1007/s00299-010-0884-y
|
| [30] |
ZHOU S, SAUVE R, HOWARD E. Identification of a cell wall peroxidase in red calli of Prunus incisa Thunb. Plant Cell Reports, 2002, 21(4):380-384.
doi: 10.1007/s00299-002-0527-z
|
| [31] |
REICHEL M, CARLE R, SRUAMSIRI P, NEIDHART S. Changes in flavonoids and nonphenolic pigments during on-tree maturation and postharvest pericarp browning of litchi (Litchi chinensis Sonn.) as shown by HPLC-MSn. Journal of Agricultural and Food Chemistry, 2011, 59:3924-3939.
doi: 10.1021/jf104432r
|
| [32] |
FANG F, ZHANG X L, LUO H H, ZHOU J J, GONG Y H, LI W J, SHI Z W, HE Q A, WU Q, LI L, JIANG L L, CAI Z G, OREN-SHAMIR M, ZHANG Z Q, PANG X Q. An intracellular laccase is responsible for epicatechin-mediated anthocyanin degradation in litchi fruit pericarp. Plant Physiology, 2015, 169(4):2391-2408.
|







 ),孙冷雪2,郑嘉敏2,蔡灿军2,王蓓2,李开拓2,潘腾飞1,2,佘文琴1,2(
),孙冷雪2,郑嘉敏2,蔡灿军2,王蓓2,李开拓2,潘腾飞1,2,佘文琴1,2(