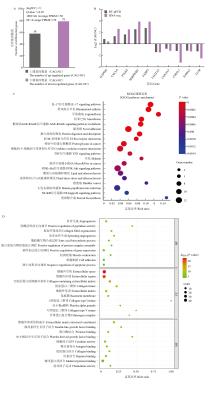
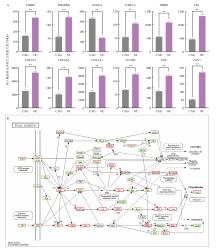
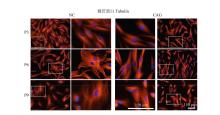
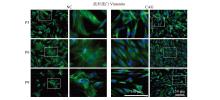
| [1] |
ZHAO Q-B, LÓPEZ-CORTEGANO E, OYELAMI F O, ZHANG Z, MA P-P, WANG Q-S, PAN Y-C. Conservation priorities analysis of Chinese indigenous pig breeds in the Taihu Lake Region. Frontiers in Genetics, 2021, 12: 558873.
|
| [2] |
孟欣. 我国猪的地方品种保护现状及对策. 中国畜牧业, 2023(17): 50-51.
|
|
MENG X. Current situation and countermeasures of local pig breeds protection in China. China Animal Industry, 2023(17): 50-51. (in Chinese)
|
| [3] |
方晓敏, 顾岳清, 黄媛, 李顺. 二花脸猪种质特性发展现状及保种建议. 中国畜禽种业, 2023, 19(8): 53-57.
|
|
FANG X M, GU Y Q, HUANG Y, LI S. Development status and conservation suggestions of Erhualian pig germplasm characteristics. The Chinese Livestock and Poultry Breeding, 2023, 19(8): 53-57. (in Chinese)
|
| [4] |
郭建凤, 孙延晓, 赵雪艳, 魏述东, 武英. 不同屠宰体重莱芜猪胴体性能及肉品质比较. 养猪, 2022(4): 48-49.
|
|
GUO J F, SUN Y X, ZHAO X Y, WEI S D, WU Y. Comparison of carcass performance and meat quality of Laiwu pigs with different slaughter weights. Swine Production, 2022(4): 48-49. (in Chinese)
|
| [5] |
苏正成, 王伟儒, 俸祥仁, 咸燕燕, 甘庆宾, 项凯琳, 兰群丽, 黄甫彦, 韦巧燕, 陆晶山. 巴马香猪资源的开发利用前景与展望. 畜牧业环境, 2024(3): 7-8.
|
|
SU Z C, WANG W R, FENG X R, XIAN Y Y, GAN Q B, XIANG K L, LAN Q L, HUANG F Y, WEI Q Y, LU J S. Development and utilization prospect of Bama Xiang pig resources. Animal Industry and Environment, 2024(3): 7-8. (in Chinese)
|
| [6] |
BIERMANN A D M, YIN T, KÖNIG VON BORSTEL U U, KRÜBESAM K, KUHN B, KÖNIG S. From phenotyping towards breeding strategies: using in vivo indicator traits and genetic markers to improve meat quality in an endangered pig breed. Animal, 2015, 9(6): 919-927.
|
| [7] |
LI L, REN Z W, WANG Q H, GE S Q, LIU Y T, LIU C J, LIU F X, HU Y X, LI J M, BAO J Y, et al. Infection of African swine fever in wild boar, China, 2018 . Transboundary and Emerging Diseases, 2019, 66(3): 1395-1398.
|
| [8] |
BORGES A A, PEREIRA A F. Potential role of intraspecific and interspecific cloning in the conservation of wild mammals. Zygote, 2019, 27(3): 111-117.
doi: 10.1017/S0967199419000170
pmid: 31182179
|
| [9] |
IQBAL A, PING J, ALI S, ZHEN G, KANG J Z, YI P Z, LU H X, ZHAO Z H. Conservation of endangered species through somatic cell nuclear transfer (SCNT). Conservation Genetics Resources, 2021, 13(3): 349-357.
|
| [10] |
GAVIN-PLAGNE L, PEROLD F, OSTEIL P, VOISIN S, MOREIRA S C, COMBOURIEU Q, SAÏDOU V, MURE M, LOUIS G, BAUDOT A, et al. Insights into species preservation: Cryobanking of rabbit somatic and pluripotent stem cells. International Journal of Molecular Sciences, 2020, 21(19): 7285.
|
| [11] |
TIAN X C, KUBOTA C, ENRIGHT B, YANG X Z. Cloning animals by somatic cell nuclear transfer: biological factors. Reproductive Biology and Endocrinology, 2003, 1: 98.
|
| [12] |
GUO Z H, LV L, LIU D, FU B. Effects of trichostatin A on pig SCNT blastocyst formation rate and cell number: a meta-analysis. Research in Veterinary Science, 2018, 117: 161-166.
doi: S0034-5288(17)31139-6
pmid: 29277014
|
| [13] |
GOUVEIA C, HUYSER C, EGLI D, PEPPER M S. Lessons learned from somatic cell nuclear transfer. International Journal of Molecular Sciences, 2020, 21(7): 2314.
|
| [14] |
KIM E, HYUN S H. Comparative analysis of various donor cell types for somatic cell nuclear transfer and its association with apoptosis and senescence. Molecular Medicine Reports, 2014, 9(1): 63-68.
doi: 10.3892/mmr.2013.1776
pmid: 24190282
|
| [15] |
LI Z C, SHI J S, LIU D W, ZHOU R, ZENG H Y, ZHOU X, MAI R B, ZENG S F, LUO L H, YU W X, et al. Effects of donor fibroblast cell type and transferred cloned embryo number on the efficiency of pig cloning. Cellular Reprogramming, 2013, 15(1): 35-42.
doi: 10.1089/cell.2012.0042
pmid: 23256540
|
| [16] |
OH H J, PARK J E, PARK E J, KIM M J, KIM G A, RHEE S H, LIM S H, KANG S K, LEE B C. Analysis of cell growth and gene expression of porcine adipose tissue-derived mesenchymal stem cells as nuclear donor cell. Development, Growth & Differentiation, 2014, 56(9): 595-604.
|
| [17] |
ZHAI Y H, LI W, ZHANG Z R, CAO Y Q, WANG Z Z, ZHANG S, LI Z Y. Epigenetic states of donor cells significantly affect the development of somatic cell nuclear transfer (SCNT) embryos in pigs. Molecular Reproduction and Development, 2018, 85(1): 26-37.
doi: 10.1002/mrd.22935
pmid: 29205617
|
| [18] |
LEE J, LEE Y J, LEE G S, LEE S T, LEE E. Comparative study of the developmental competence of cloned pig embryos derived from spermatogonial stem cells and fetal fibroblasts. Reproduction in Domestic Animals, 2019, 54(9): 1258-1264.
doi: 10.1111/rda.13507
pmid: 31283039
|
| [19] |
SHARMA P, YADAV A S, SELOKAR N L, KUMAR D, DHAKA S S, YADAV P S. Epigenetic status of buffalo fibroblasts treated with sodium butyrate a chromatin remodeling agent. Tissue and Cell, 2018, 50: 51-58.
|
| [20] |
ZHU Y K, LIU X W, DING X L, WANG F, GENG X. Telomere and its role in the aging pathways: Telomere shortening, cell senescence and mitochondria dysfunction. Biogerontology, 2019, 20(1): 1-16.
doi: 10.1007/s10522-018-9769-1
pmid: 30229407
|
| [21] |
OSBALDISTON G W, COFFMAN J R, STOWE E C. Equine isoerythrolysis: clinical pathological observations and transfusion of dam’s red blood cells to her foal. Canadian Journal of Comparative Medicine, 1969, 33(4): 310-315.
|
| [22] |
许卫华, 李紫聪, 吴珍芳, 石俊松. DNMT1基因干扰对猪体细胞克隆胚胎发育、基因转录和DNA甲基化的影响. 农业生物技术学报, 2018, 26(12): 2075-2083.
|
|
XU W H, LI Z C, WU Z F, SHI J S. Effect of DNMT1 interference on development, gene transcription and DNA methylation of porcine(sus scrofa) cloned embryos. Journal of Agricultural Biotechnology, 2018, 26(12): 2075-2083. (in Chinese)
|
| [23] |
唐红, 张宾, 张译元, 郭延华, 王立民, 江中良, 周平. 供体细胞对绵羊转基因体细胞克隆胚胎体外发育影响的研究. 家畜生态学报, 2016, 37(10): 16-21.
|
|
TANG H, ZHANG B, ZHANG Y Y, GUO Y H, WANG L M, JIANG Z L, ZHOU P. Effect of donor cell treatment on the development in vitro of reconstructed transgenic sheep embryos. Journal of Domestic Animal Ecology, 2016, 37(10): 16-21. (in Chinese)
|
| [24] |
MUÑOZ-ESPÍN D, SERRANO M. Cellular senescence: From physiology to pathology. Nature Reviews Molecular Cell Biology, 2014, 15(7): 482-496.
|
| [25] |
NELSON G, WORDSWORTH J, WANG C F, JURK D, LAWLESS C, MARTIN-RUIZ C, VON ZGLINICKI T. A senescent cell bystander effect: senescence-induced senescence. Aging Cell, 2012, 11(2): 345-349.
doi: 10.1111/j.1474-9726.2012.00795.x
pmid: 22321662
|
| [26] |
WLASCHEK M, MAITY P, MAKRANTONAKI E, SCHARFFETTER- KOCHANEK K. Connective tissue and fibroblast senescence in skin aging. Journal of Investigative Dermatology, 2021, 141(4): 985-992.
|
| [27] |
KASPER G, MAO L, GEISSLER S, DRAYCHEVA A, TRIPPENS J, KÜHNISCH J, TSCHIRSCHMANN M, KASPAR K, PERKA C, DUDA G N, KLOSE J. Insights into mesenchymal stem cell aging: Involvement of antioxidant defense and actin cytoskeleton. Stem Cells, 2009, 27(6): 1288-1297.
doi: 10.1002/stem.49
pmid: 19492299
|
| [28] |
SAMIEC M, SKRZYSZOWSKA M. Extranuclear inheritance of mitochondrial genome and epigenetic reprogrammability of chromosomal telomeres in somatic cell cloning of mammals. International Journal of Molecular Sciences, 2021, 22(6): 3099.
|
| [29] |
GIANCOTTI F G, RUOSLAHTI E. Integrin signaling. Science, 1999, 285(5430): 1028-1033.
doi: 10.1126/science.285.5430.1028
pmid: 10446041
|
| [30] |
MU X D, TSENG C, HAMBRIGHT W S, MATRE P, LIN C Y, CHANDA P, CHEN W Q, GU J H, RAVURI S, CUI Y, et al. Cytoskeleton stiffness regulates cellular senescence and innate immune response in Hutchinson-Gilford Progeria Syndrome. Aging Cell, 2020, 19(8): e13152.
|
| [31] |
D’ANGELO M, BENEDETTI E, TUPONE M G, CATANESI M, CASTELLI V, ANTONOSANTE A, CIMINI A. The role of stiffness in cell reprogramming: A potential role for biomaterials in inducing tissue regeneration. Cells, 2019, 8(9): 1036.
|
| [32] |
CHEN X, TANG W J, SHI J B, LIU M M, LIU X H. Therapeutic strategies for targeting telomerase in cancer. Medicinal Research Reviews, 2020, 40(2): 532-585.
doi: 10.1002/med.21626
pmid: 31361345
|
| [33] |
YANG M H, HWANG S T, UM J Y, AHN K S. Cycloastragenol exerts protective effects against UVB irradiation in human dermal fibroblasts and HaCaT keratinocytes. Journal of Dermatological Science, 2023, 111(2): 60-67.
doi: 10.1016/j.jdermsci.2023.07.001
pmid: 37474410
|
| [34] |
IKRAM M, JO M H, CHOE K, KHAN A, AHMAD S, SAEED K, KIM M W, KIM M O. Cycloastragenol, a triterpenoid saponin, regulates oxidative stress, neurotrophic dysfunctions, neuroinflammation and apoptotic cell death in neurodegenerative conditions. Cells, 2021, 10(10): 2719.
|
| [35] |
ULLAH M, SUN Z J. Klotho deficiency accelerates stem cells aging by impairing telomerase activity. The Journals of Gerontology Series A, Biological Sciences and Medical Sciences, 2019, 74(9): 1396-1407.
|
| [36] |
IP F C F, NG Y P, AN H J, DAI Y, PANG H H, HU Y Q, CHIN A C, HARLEY C B, WONG Y H, IP N Y. Cycloastragenol is a potent telomerase activator in neuronal cells: implications for depression management. Neuro-Signals, 2014, 22(1): 52-63.
|
| [37] |
WU J H, ZENG Z W, LI Y Y, QIN H Y, ZUO C Q, ZHOU C H, XU D H. Cycloastragenol protects against glucocorticoid-induced osteogenic differentiation inhibition by activating telomerase. Phytotherapy Research, 2021, 35(4): 2034-2044.
doi: 10.1002/ptr.6946
pmid: 33165990
|
| [38] |
ZHANG J, HAO L L, WEI Q, ZHANG S, CHENG H, ZHAI Y H, JIANG Y, AN X L, LI Z Y, ZHANG X M, TANG B. TET 3 overexpression facilitates DNA reprogramming and early development of bovine SCNT embryos. Reproduction, 2020, 160(3): 379-391.
|
| [39] |
SWEGEN A, APPELTANT R, WILLIAMS S A. Cloning in action: Can embryo splitting, induced pluripotency and somatic cell nuclear transfer contribute to endangered species conservation? Biological Reviews, 2023, 98(4): 1225-1249.
|
| [40] |
RODIER F, CAMPISI J. Four faces of cellular senescence. The Journal of Cell Biology, 2011, 192(4): 547-556.
|
| [41] |
ZHANG Y H, GAO D X, YUAN Y, ZHENG R Z, SUN M T, JIA S T, LIU J. Cycloastragenol: a novel senolytic agent that induces senescent cell apoptosis and restores physical function in TBI-aged mice. International Journal of Molecular Sciences, 2023, 24(7): 6554.
|
| [42] |
DONG Z N, LUO Y H, YUAN Z C, TIAN Y, JIN T Q, XU F. Cellular senescence and SASP in tumor progression and therapeutic opportunities. Molecular Cancer, 2024, 23(1): 181.
doi: 10.1186/s12943-024-02096-7
pmid: 39217404
|
| [43] |
ALESSIO N, SQUILLARO T, DI BERNARDO G, GALANO G, DE ROSA R, MELONE M A B, PELUSO G, GALDERISI U. Increase of circulating IGFBP-4 following genotoxic stress and its implication for senescence. eLife, 2020, 9: e54523.
|
| [44] |
MERCURIO L, LULLI D, MASCIA F, DELLAMBRA E, SCARPONI C, MORELLI M, VALENTE C, CARBONE M L, PALLOTTA S, GIROLOMONI G, et al. Intracellular Insulin-like growth factor binding protein 2 (IGFBP2) contributes to the senescence of keratinocytes in psoriasis by stabilizing cytoplasmic p21. Aging, 2020, 12(8): 6823-6851.
|
| [45] |
CUI N, HU M, KHALIL R A. Chapter one biochemical and biological attributes of matrix metalloproteinases. Progress in Molecular Biology and Translational Science, 2017, 147: 1-73.
doi: S1877-1173(17)30032-7
pmid: 28413025
|
| [46] |
CABRAL-PACHECO G A, GARZA-VELOZ I, CASTRUITA-DE LA ROSA C, RAMIREZ-ACUÑA J M, PEREZ-ROMERO B A, GUERRERO-RODRIGUEZ J F, MARTINEZ-AVILA N, MARTINEZ- FIERRO M L. The roles of matrix metalloproteinases and their inhibitors in human diseases. International Journal of Molecular Sciences, 2020, 21(24): 9739.
|
| [47] |
GUTIÉRREZ-FERNÁNDEZ A, SORIA-VALLES C, OSORIO F G, GUTIÉRREZ-ABRIL J, GARABAYA C, AGUIRRE A, FUEYO A, FERNÁNDEZ-GARCÍA M S, PUENTE X S, LÓPEZ-OTÍN C. Loss of MT1-MMP causes cell senescence and nuclear defects which can be reversed by retinoic acid. The EMBO Journal, 2015, 34(14): 1875-1888.
|
| [48] |
HASSONA Y, CIRILLO N, HEESOM K, PARKINSON E K, PRIME S S. Senescent cancer-associated fibroblasts secrete active MMP-2 that promotes keratinocyte dis-cohesion and invasion. British Journal of Cancer, 2014, 111(6): 1230-1237.
doi: 10.1038/bjc.2014.438
pmid: 25117810
|
| [49] |
LEVI N, PAPISMADOV N, SOLOMONOV I, SAGI I, KRIZHANOVSKY V. The ECM path of senescence in aging: components and modifiers. The FEBS Journal, 2020, 287(13): 2636-2646.
|
| [50] |
LIU J Q, XIAO Q, XIAO J N, NIU C X, LI Y Y, ZHANG X J, ZHOU Z W, SHU G, YIN G. Wnt/β-catenin signalling: function, biological mechanisms, and therapeutic opportunities. Signal Transduction and Targeted Therapy, 2022, 7(1): 3.
doi: 10.1038/s41392-021-00762-6
pmid: 34980884
|
| [51] |
MIRZOEV T M, SHARLO K A, SHENKMAN B S. The role of GSK-3β in the regulation of protein turnover, myosin phenotype, and oxidative capacity in skeletal muscle under disuse conditions. International Journal of Molecular Sciences, 2021, 22(10): 5081.
|
| [52] |
NAKAJIMA K, OKA S, TANIKAWA T, NEMOTO-SASAKI Y, MATSUMOTO N, ISHIGURO H, ARATA Y, SUGIURA T, YAMASHITA A. Lysophosphatidylinositol induced morphological changes and stress fiber formation through the GPR55-RhoA-ROCK pathway. International Journal of Molecular Sciences, 2022, 23(18): 10932.
|
| [53] |
MACHESKY L M. CYRI proteins: controllers of actin dynamics in the cellular ‘eat vs walk’ decision. Biochemical Society Transactions, 2023, 51(2): 579-585.
|
| [54] |
MU X D, LIN C Y, HAMBRIGHT W S, TANG Y, RAVURI S, LU A P, MATRE P, CHEN W Q, GAO X Q, CUI Y, et al. Aberrant RhoA activation in macrophages increases senescence-associated secretory phenotypes and ectopic calcification in muscular dystrophic mice. Aging, 2020, 12(24): 24853-24871.
|
| [55] |
NING L, GAO L, ZHANG F, LI X X, WANG T T. Mechanical stretch induces annulus fibrosus cell senescence through activation of the RhoA/ROCK pathway. BioMed Research International, 2021, 2021(1): 5321121.
|
| [56] |
HUMPHRIES J D, WANG P B, STREULI C, GEIGER B, HUMPHRIES M J, BALLESTREM C. Vinculin controls focal adhesion formation by direct interactions with talin and actin. The Journal of Cell Biology, 2007, 179(5): 1043-1057.
|
| [57] |
MISHRA Y G, MANAVATHI B. Focal adhesion dynamics in cellular function and disease. Cellular Signalling, 2021, 85: 110046.
|
| [58] |
POLLARD T D, COOPER J A. Actin, a central player in cell shape and movement. Science, 2009, 326(5957): 1208-1212.
doi: 10.1126/science.1175862
pmid: 19965462
|
| [59] |
KITAZAWA T, SHUMAN H, SOMLYO A P. Calcium and magnesium binding to thin and thick filaments in skinned muscle fibres: Electron probe analysis. Journal of Muscle Research and Cell Motility, 1982, 3(4): 437-454.
pmid: 6985147
|
| [60] |
WANG E, GUNDERSEN D. Increased organization of cytoskeleton accompanying the aging of human fibroblasts in vitro. Experimental Cell Research, 1984, 154(1): 191-202.
|
| [61] |
MCBEATH R, PIRONE D M, NELSON C M, BHADRIRAJU K, CHEN C S. Cell shape, cytoskeletal tension, and RhoA regulate stem cell lineage commitment. Developmental Cell, 2004, 6(4): 483-495.
doi: 10.1016/s1534-5807(04)00075-9
pmid: 15068789
|
| [62] |
SLIOGERYTE K, GAVARA N. Vimentin plays a crucial role in fibroblast ageing by regulating biophysical properties and cell migration. Cells, 2019, 8(10): 1164.
|
| [63] |
STANULIS-PRAEGERS B M, YAAR M, GILCHREST B A. Evidence that filopodia outgrowth is a common final pathway for fibroblast growth inhibition in vitro. Experimental Dermatology, 1992, 1(3): 141-148.
|
| [64] |
KUMAR S, MAXWELL I Z, HEISTERKAMP A, POLTE T R, LELE T P, SALANGA M, MAZUR E, INGBER D E. Viscoelastic retraction of single living stress fibers and its impact on cell shape, cytoskeletal organization, and extracellular matrix mechanics. Biophysical Journal, 2006, 90(10): 3762-3773.
pmid: 16500961
|
| [65] |
VEITIA R A. DNA content, cell size, and cell senescence. Trends in Biochemical Sciences, 2019, 44(8): 645-647.
doi: S0968-0004(19)30107-0
pmid: 31160123
|
| [66] |
RIEGGER J, BRENNER R E. Increase of cell surface vimentin is associated with vimentin network disruption and subsequent stress-induced premature senescence in human chondrocytes. eLife, 2023, 12: e91453.
|
| [67] |
NISHIO K, INOUE A, QIAO S L, KONDO H, MIMURA A. Senescence and cytoskeleton: Overproduction of vimentin induces senescent-like morphology in human fibroblasts. Histochemistry and Cell Biology, 2001, 116(4): 321-327.
pmid: 11702190
|
| [68] |
WHITWORTH K M, PRATHER R S. Somatic cell nuclear transfer efficiency: How can it be improved through nuclear remodeling and reprogramming? Molecular Reproduction and Development, 2010, 77(12): 1001-1015.
doi: 10.1002/mrd.21242
pmid: 20931660
|
| [69] |
NADRI P, ANSARI-MAHYARI S, JAFARPOUR F, MAHDAVI A H, TANHAEI VASH N, LACHINANI L, DORMIANI K, NASR-ESFAHANI M H. Melatonin accelerates the developmental competence and telomere elongation in ovine SCNT embryos. PLoS ONE, 2022, 17(7): e0267598.
|
| [70] |
ZHANG H D, WU B, LIU H L, QIU M N, LIU J, ZHANG Y, QUAN F S. Improving development of cloned goat embryos by supplementing α-lipoic acid to oocyte in vitro maturation medium. Theriogenology, 2013, 80(3): 228-233.
|
| [71] |
SRIRATTANA K, KANEDA M, PARNPAI R. Strategies to improve the efficiency of somatic cell nuclear transfer. International Journal of Molecular Sciences, 2022, 23(4): 1969.
|
| [72] |
SONG Y R, HAI T, WANG Y, GUO R F, LI W, WANG L, ZHOU Q. Epigenetic reprogramming, gene expression and in vitro development of porcine SCNT embryos are significantly improved by a histone deacetylase inhibitor: m-carboxycinnamic acid bishydroxamide (CBHA). Protein & Cell, 2014, 5(5): 382-393.
|
| [73] |
ZHANG S, CHEN X, WANG F, AN X L, TANG B, ZHANG X M, SUN L G, LI Z Y. Aberrant DNA methylation reprogramming in bovine SCNT preimplantation embryos. Scientific Reports, 2016, 6: 30345.
doi: 10.1038/srep30345
pmid: 27456302
|
| [74] |
ASHRY M, YANG C Y, RAJPUT S K, FOLGER J K, KNOTT J G, SMITH G W. Follistatin supplementation induces changes in CDX2 CpG methylation and improves in vitro development of bovine SCNT preimplantation embryos. Reproductive Biology and Endocrinology, 2021, 19(1): 141.
|
| [75] |
KOHAN-GHADR H R, SMITH L C, ARNOLD D R, MURPHY B D, LEFEBVRE R C. Aberrant expression of E-cadherin and β-catenin proteins in placenta of bovine embryos derived from somatic cell nuclear transfer. Reproduction, Fertility, and Development, 2012, 24(4): 588-598.
|
| [76] |
BURGSTALLER J P, BREM G. Aging of cloned animals: A mini-review. Gerontology, 2017, 63(5): 417-425.
doi: 10.1159/000452444
pmid: 27820924
|
| [77] |
WANG Y X, CHEN C, WANG Q Y, CAO Y N, XU L, QI R. Inhibitory effects of cycloastragenol on abdominal aortic aneurysm and its related mechanisms. British Journal of Pharmacology, 2019, 176(2): 282-296.
doi: 10.1111/bph.14515
pmid: 30302749
|







 ), ZHANG Liang1,2(
), ZHANG Liang1,2(