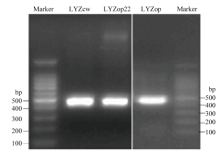
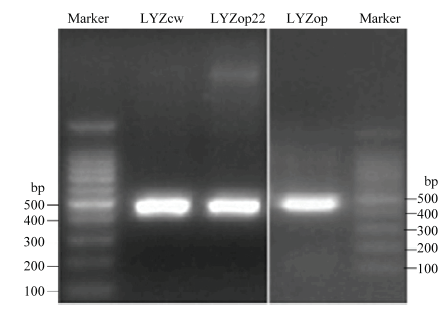
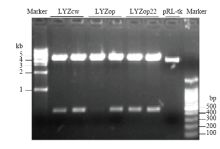
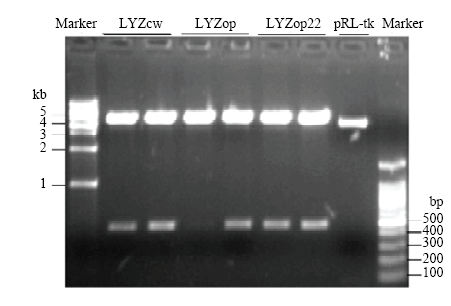
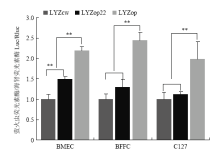
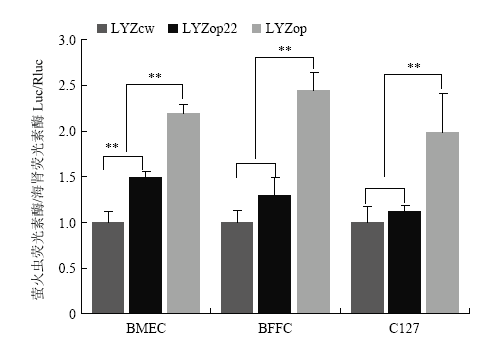
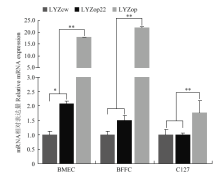
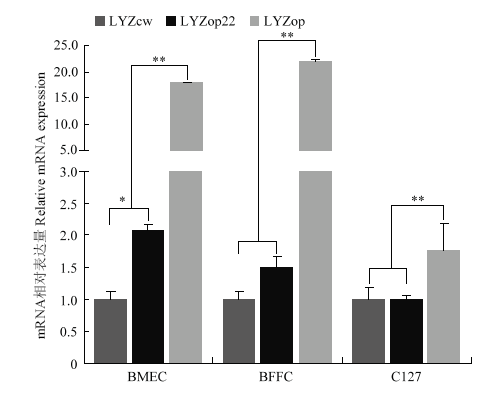
Scientia Agricultura Sinica ›› 2020, Vol. 53 ›› Issue (18): 3805-3817.doi: 10.3864/j.issn.0578-1752.2020.18.015
• ANIMAL SCIENCE·VETERINARY SCIENCE·RESOURCE INSECT • Previous Articles Next Articles
TIAN Yuan1( ),WANG Li1,LONG Feng1,ZAN LinSen1,2,CHENG Gong1,2()
),WANG Li1,LONG Feng1,ZAN LinSen1,2,CHENG Gong1,2()
| [1] | 李永富, 孙震, 史锋, 张颖, 马金生, 蒋立凤. 溶菌酶对猪肉的保鲜作用. 上海农业学报, 2009,25(04):61-63. |
| LI Y F, SUN Z, SHI F, ZHANG Y, MA J S, JIANG L F. The effect of lysozyme on keeping chilled pork fresh. Acta Agriculturae Shanghai, 2009,25(04):61-63. (in Chinese) | |
| [2] | 朱军英, 崔立, 沈彦萍, 施彬. 溶菌酶对仔猪腹泻的预防作用. 中国畜牧杂志, 2009,45(03):40-42. |
| ZHU J Y, CUI L, SHEN Y P, SHI B. The preventive effect of lysozyme on diarrhea in piglets. Chinese Journal of Animal Science, 2009,45(03):40-42. (in Chinese) | |
| [3] |
RAGLAND S A, CRISS A K. From bacterial killing to immune modulation: Recent insights into the functions of lysozyme. PLoS Pathogens, 2017,13(9):e1006512.
doi: 10.1371/journal.ppat.1006512 pmid: 28934357 |
| [4] |
THELLIN O, ZORZI D, MELIN P, DELVENNE P, HEINEN E, ZORZI W, EL MOUALIJ B, QUATRESOOZ P. Hen egg white lysozyme is less active against Gardnerella vaginalis biofilm than human lysozyme. Microbiology Research International, 2018,6(1):1-6.
doi: 10.30918/MRI |
| [5] |
ERCAN D, DEMIRCI A. Recent advances for the production and recovery methods of lysozyme. Critical Reviews in Biotechnology, 2016,36(6):1078-1088.
doi: 10.3109/07388551.2015.1084263 pmid: 26383819 |
| [6] |
MAGA E A, SHOEMAKER C F, ROWE J D, BONDURANT R H, ANDERSON G B, MURRAY J D. Production and processing of milk from transgenic goats expressing human lysozyme in the mammary gland. Journal of Dairy Science, 2006,89(2):518-524.
doi: 10.3168/jds.S0022-0302(06)72114-2 pmid: 16428620 |
| [7] |
LIU X, WANG Y, TIAN Y, YU Y, GAO M, HU G, SU F, PAN S, LUO Y, GUO Z, QUAN F, ZHANG Y. Generation of mastitis resistance in cows by targeting human lysozyme gene to beta-casein locus using zinc-finger nucleases. Proceedings of the Royal Society B Biological Sciences, 2014,281(1780):20133368.
doi: 10.1098/rspb.2013.3368 |
| [8] | 熊显荣, 李文哲, 王丽君, 王勇胜, 苏建民, 华松, 张涌. 体细胞核移植技术生产转人溶菌酶基因(hLYZ)牛胚胎的研究. 农业生物技术学报, 2011,19(02):294-301. |
| XIONG X R, LI W Z, WANG L J, WANG Y S, SU J M, HUA S, ZHANG Y. Human lysozyme gene (hLYZ) transgenic bovine embryos produced by somatic cell nuclear transfer. Journal of Agricultural Biotechnology, 2011,19(02):294-301. (in Chinese) | |
| [9] |
YANG B, WANG J, TANG B, LIU Y, GUO C, YANG P, YU T, LI R, ZHAO J, ZHANG L, DAI Y, LI N. Characterization of bioactive recombinant human lysozyme expressed in milk of cloned transgenic cattle. PLoS One, 2011,6(3):e17593.
doi: 10.1371/journal.pone.0017593 pmid: 21436886 |
| [10] |
LU D, LIU S, SHANG S, WU F, WEN X, LI Z, LI Y, HU X, ZHAO Y, LI Q, LI N. Production of transgenic-cloned pigs expressing large quantities of recombinant human lysozyme in milk. PLoS One, 2015,10(5):e0123551.
doi: 10.1371/journal.pone.0123551 pmid: 25955256 |
| [11] |
YU H, CHEN J, LIU S, ZHANG A, XU X, WANG X, LU P, CHENG G. Large-scale production of functional human lysozyme in transgenic cloned goats. Journal of Biotechnology, 2013,168(4):676-683.
doi: 10.1016/j.jbiotec.2013.10.023 |
| [12] |
MAURO V P, CHAPPELL S A. Considerations in the use of codon optimization for recombinant protein expression. Methods in Molecular Biology, 2018,1850:275-288.
doi: 10.1007/978-1-4939-9086-3_20 pmid: 30788833 |
| [13] |
LI G W, OH E, WEISSMAN J S. The anti-Shine-Dalgarno sequence drives translational pausing and codon choice in bacteria. Nature, 2012,484(7395):538-541.
doi: 10.1038/nature10965 |
| [14] |
HANSON G, COLLER J. Codon optimality, bias and usage in translation and mRNA decay. Nature Reviews Molecular Cell Biology, 2018,19(1):20-30.
doi: 10.1038/nrm.2017.91 pmid: 29018283 |
| [15] |
CARNEIRO R L, REQUIAO R D, ROSSETTO S, DOMITROVIC T, PALHANO F L. Codon stabilization coefficient as a metric to gain insights into mRNA stability and codon bias and their relationships with translation. Nucleic Acids Research, 2019,47(5):2216-2228.
doi: 10.1093/nar/gkz033 pmid: 30698781 |
| [16] |
GUSTAFSSON C, GOVINDARAJAN S, MINSHULL J. Codon bias and heterologous protein expression. Trends in Biotechnology, 2004,22(7):346-353.
doi: 10.1016/j.tibtech.2004.04.006 |
| [17] |
COOPER C A, MAGA E A, MURRAY J D. Production of human lactoferrin and lysozyme in the milk of transgenic dairy animals: past, present, and future. Transgenic Research, 2015,24(4):605-614.
doi: 10.1007/s11248-015-9885-5 pmid: 26059245 |
| [18] |
PLOTKIN J B, ROBINS H, LEVINE A J. Tissue-specific codon usage and the expression of human genes. Proceedings of the National Academy of Sciences of the United States of America, 2004,101(34):12588-12591.
doi: 10.1073/pnas.0404957101 pmid: 15314228 |
| [19] |
NEWMAN Z R, YOUNG J M, INGOLIA N T, BARTON G M. Differences in codon bias and GC content contribute to the balanced expression of TLR7 and TLR9. Proceedings of the National Academy of Sciences, 2016,113(10):e1362-e1371.
doi: 10.1073/pnas.1518976113 |
| [20] |
KUDLA G, LIPINSKI L, CAFFIN F, HELWAK A, ZYLICZ M. High guanine and cytosine content increases mRNA levels in mammalian cells. PLoS Biology, 2006,4(6):e180.
doi: 10.1371/journal.pbio.0040180 pmid: 16700628 |
| [21] |
PLOTKIN J B, KUDLA G. Synonymous but not the same: the causes and consequences of codon bias. Nature Reviews Genetics, 2011,12(1):32-42.
doi: 10.1038/nrg2899 pmid: 21102527 |
| [22] |
BOEL G, LETSO R, NEELY H, PRICE W N, WONG K H, SU M, LUFF J, VALECHA M, EVERETT J K, ACTON T B, XIAO R, MONTELIONE G T, AALBERTS D P, HUNT J F. Codon influence on protein expression in E. coli correlates with mRNA levels. Nature, 2016,529(7586):358-363.
doi: 10.1038/nature16509 pmid: 26760206 |
| [23] |
HANSON G, COLLER J. Codon optimality, bias and usage in translation and mRNA decay. Nature Reviews Molecular Cell Biology, 2018,19(1):20-30.
doi: 10.1038/nrm.2017.91 pmid: 29018283 |
| [24] | WANG X, LI P, GUTENKUNST R N. Systematic effects of mRNA secondary structure on gene expression and molecular function in budding yeast. BioRxiv, 2017: 138792. |
| [25] |
ZUR H, TULLER T. Strong association between mRNA folding strength and protein abundance in S. cerevisiae. Embo Reports, 2012,13(3):272-277.
doi: 10.1038/embor.2011.262 |
| [26] |
JAGODNIK J, CHIARUTTINI C, GUILLIER M. Stem-loop structures within mRNA coding sequences activate translation initiation and mediate control by small regulatory RNAs. Molecular Cell, 2017, 68(1): 158-170.e3.
doi: 10.1016/j.molcel.2017.08.015 pmid: 28918899 |
| [27] |
TULLER T, ZUR H. Multiple roles of the coding sequence 5' end in gene expression regulation. Nucleic Acids Research, 2015,43(1):13-28.
doi: 10.1093/nar/gku1313 pmid: 25505165 |
| [28] |
PAULUS M, HASLBECK M, WATZELE M. RNA stem-loop enhanced expression of previously non-expressible genes. Nucleic Acids Research, 2004,32(9):e78.
doi: 10.1093/nar/gnh076 pmid: 15163763 |
| [29] |
MAO Y, LIU H, LIU Y, TAO S. Deciphering the rules by which dynamics of mRNA secondary structure affect translation efficiency in Saccharomyces cerevisiae. Nucleic Acids Research, 2014,42(8):4813-4822.
doi: 10.1093/nar/gku159 pmid: 24561808 |
| [30] |
ZHOU Z, DANG Y, ZHOU M, LI L, YU C, FU J, CHEN S, LIU Y. Codon usage is an important determinant of gene expression levels largely through its effects on transcription. Proceedings of the National Academy of Sciences, 2016,113(41):e6117.
doi: 10.1073/pnas.1606724113 |
| [31] |
PRESNYAK V, ALHUSAINI N, CHEN Y H, MARTIN S, MORRIS N, KLINE N, OLSON S, WEINBERG D, BAKER K E, GRAVELEY B R, COLLER J. Codon optimality is a major determinant of mRNA stability. Cell, 2015,160(6):1111-1124.
doi: 10.1016/j.cell.2015.02.029 pmid: 25768907 |
| [32] |
MITARAI N, SNEPPEN K, PEDERSEN S. Ribosome collisions and translation efficiency: optimization by codon usage and mRNA destabilization. Journal of Molecular Biology, 2008,382(1):236-245.
doi: 10.1016/j.jmb.2008.06.068 pmid: 18619977 |
| [33] |
YU C H, DANG Y, ZHOU Z, WU C, ZHAO F, SACHS M S, LIU Y. Codon usage influences the local rate of translation elongation to regulate co-translational protein folding. Molecular Cell, 2015,59(5):744-754.
doi: 10.1016/j.molcel.2015.07.018 pmid: 26321254 |
| [34] |
KOUSSOUNADIS A, LANGDON S P, UM I H, HARRISON D J, SMITH V A. Relationship between differentially expressed mRNA and mRNA-protein correlations in a xenograft model system. Scientific Reports, 2015,5:10775.
doi: 10.1038/srep10775 pmid: 26053859 |
| [1] | ZHAO LiQun,QIU YanHong,ZHANG XiaoFei,LIU Hui,YANG JingJing,ZHANG Jian,ZHANG HaiJun,XU XiuLan,WEN ChangLong. The Detection of Citrullus lanatus Cryptic Virus Using TaqMan-qPCR Method [J]. Scientia Agricultura Sinica, 2021, 54(20): 4337-4347. |
| [2] | ZHAO LeiLin, FAN Xin, NIE Xing, LIANG ChengZhen, ZHANG Rui, SUN GuoQing, MENG ZhiGang, LIN Min, WANG Yuan, GUO SanDui. Salt and Drought Tolerance in Heterologous-Expression of irrE Transgenic Tobacco [J]. Scientia Agricultura Sinica, 2017, 50(20): 3860-3870. |
| [3] | . Clone and expression of broiler HSP70 gene in E. coli and preparation of HSP70 monoclonal antibody [J]. Scientia Agricultura Sinica, 2008, 41(7): 2154-2161 . |
|
||