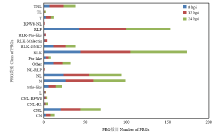
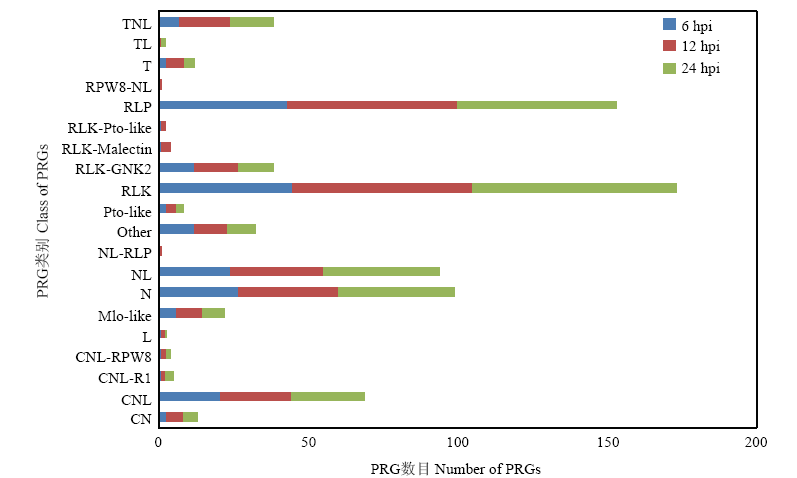
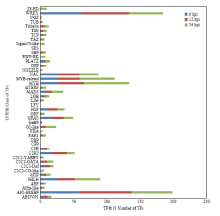
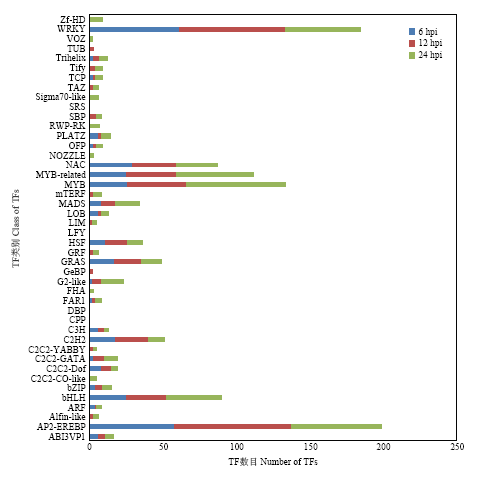
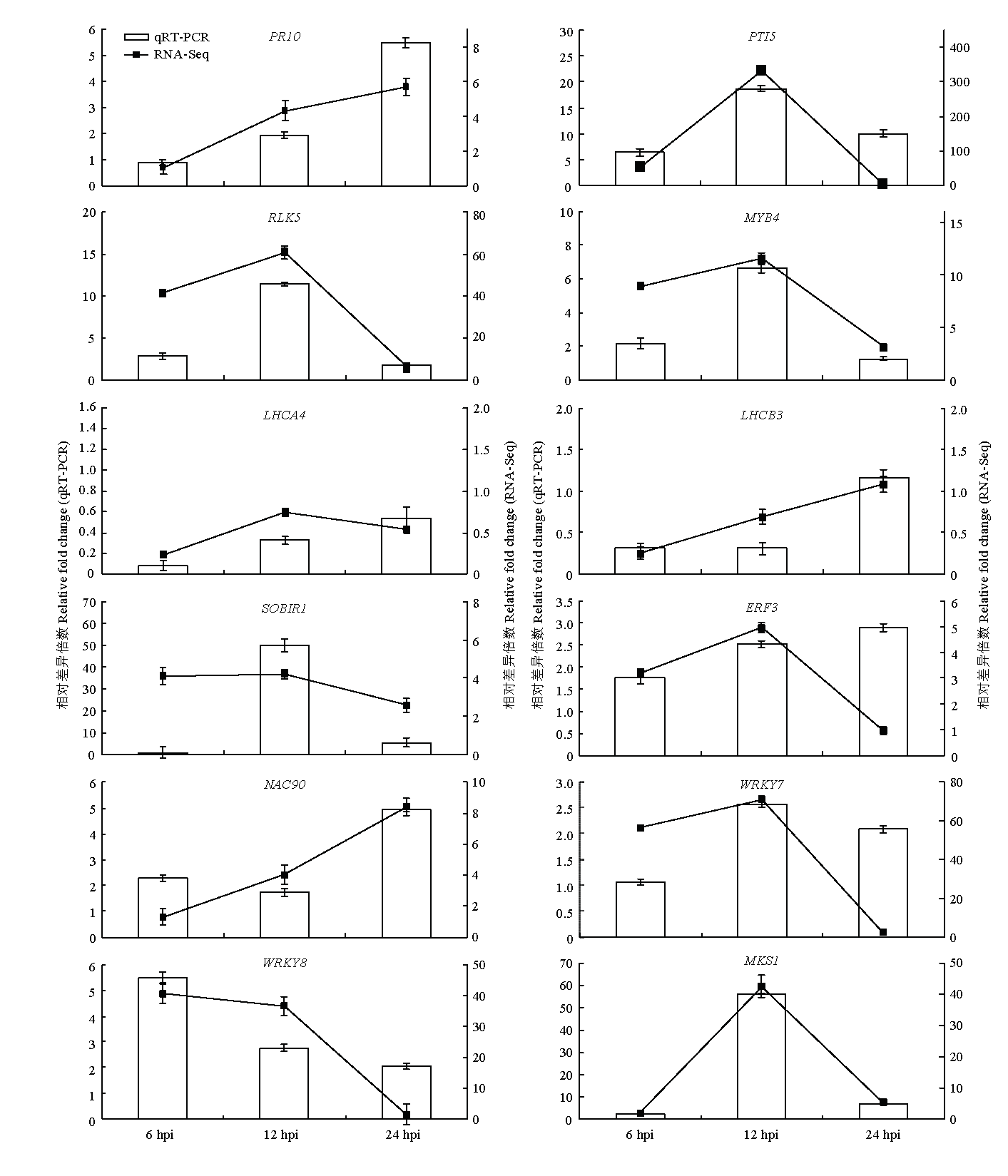
Scientia Agricultura Sinica ›› 2019, Vol. 52 ›› Issue (21): 3794-3805.doi: 10.3864/j.issn.0578-1752.2019.21.008
• PLANT PROTECTION • Previous Articles Next Articles
LIANG YingBo,LI Ze,QIU DeWen,ZENG HongMei,LI GuangYue,YANG XiuFen( )
)
| [1] | BARBARA D J, CLEWES E . Plant pathogenic Verticillium species: How many of them are there? Molecular Plant Pathology, 2003,4(4):297-305. |
| [2] | DURESSA D, ANCHIETA A, CHEN D, KLIMES A ,GARCIA-PEDRAJAS M D,DOBINSON K F,KLOSTERMAN S J. RNA-seq analyses of gene expression in the microsclerotia of Verticillium dahliae. BMC Genomics, 2013,14:607. |
| [3] | CARROLL C L, CARTER C A, GOODHUE R E ,LAWELL C C L, SUBBARAO K V. A review of control options and externalities for Verticillium wilts. Phytopathology, 2018,108(2):160-171. |
| [4] | 邱德文 . 我国植物病害生物防治的现状及发展策略. 植物保护, 2010,36(4):15-18. |
| QIU D W . Current status and development strategy for biological control of plant diseases in China. Plant Protection, 2010,36(4):15-18. (in Chinese) | |
| [5] | WANG B, YANG X, ZENG H, LIU H, ZHOU T, TAN B, YUAN J, GUO L, QIU D . The purification and characterization of a novel hypersensitive-like response-inducing elicitor from Verticillium dahliae that induces resistance responses in tobacco. Applied Microbiology and Biotechnology, 2012,93(1):191-201. |
| [6] | 王炳楠, 杨秀芬, 曾洪梅, 邱德文 . 大丽轮枝菌分泌蛋白激发子的分离纯化及生物功能研究. 生物技术通报, 2011(11):166-171. |
| WANG B N, YANG X F, ZENG H M, QIU D W . Purification and its bioassay of secreted elicitor protein fromVerticillium dahliae. Biotechnology Bulletin, 2011(11):166-171 (in Chinese) | |
| [7] | 卜冰武, 邱德文, 曾洪梅, 郭立华, 袁京京, 杨秀芬 . 大丽轮枝菌蛋白激发子PevD1诱导棉花抗病性及作用机理. 植物病理学报, 2014,44(3):254-264. |
| BU B W, QIU D W, ZENG H M, GUO L H, YUAN J J, YANG X F . Induced resistance and mechanism of protein elicitor PevD1 against Verticillium dahliae in cotton. Acta Phytopathologica Sinica, 2014,44(3):254-264. (in Chinese) | |
| [8] | ZHANG Y, GAO Y, LIANG Y, DONG Y, YANG X, QIU D . Verticillium dahliae PevD1, an Alt a 1-like protein, targets cotton PR5-like protein and promotes fungal infection. Journal of Experimental Botany, 2019,70(2):613-626. |
| [9] | WANG L K, FENG Z X, WANG X, WANG X W, ZHANG X G . DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics, 2010,26(1):136-138. |
| [10] | SCHMITTGEN T D, LEE E J, JIANG J . High-throughput real-time PCR. Methods in Molecular Biology, 2008,429:89-98. |
| [11] | SAIJO Y, LOO E P, YASUDA S . Pattern recognition receptors and signaling in plant-microbe interactions. The Plant Journal, 2018,93(4):592-613. |
| [12] | LIANG X, ZHOU J M . Receptor-like cytoplasmic kinases: Central players in plant receptor kinase-mediated signaling. Annual Review of Plant Biology, 2018,69:267-299. |
| [13] | SU J, YANG L, ZHU Q, WU H, HE Y, LIU Y, XU J, JIANG D, ZHANG S . Active photosynthetic inhibition mediated by MPK3/MPK6 is critical to effector-triggered immunity. PLoS Biology, 2018,16(5):e2004122. |
| [14] | CHINCHILLA D, ZIPFEL C, ROBATZEK S, KEMMERLING B , NÜRNBERGER T, JONES J D, FELIX G, BOLLER T. A flagellin- induced complex of the receptor FLS2 and BAK1 initiates plant defence. Nature, 2007,448(7152):497-500. |
| [15] | SCHULZE B, MENTZEL T, JEHLE A K, MUELLER K, BEELER S, BOLLER T, FELIX G, CHINCHILLA D . Rapid heteromerization and phosphorylation of ligand-activated plant transmembrane receptors and their associated kinase BAK1. The Journal of Biological Chemistry, 2010,285(13):9444-9451. |
| [16] |
LIEBRAND T W ,VAN DEN BURG H A, JOOSTEN M H. Two for all: Receptor-associated kinases SOBIR1 and BAK1. Trends in Plant Science, 2014,19(2):123-132.
doi: 10.1016/j.tplants.2013.10.003 |
| [17] | ZIPFEL C, KUNZE G, CHINCHILLA D, CANIARD A, JONES J D, BOLLER T, FELIX G . Perception of the bacterial PAMP EF-Tu by the receptor EFR restricts Agrobacterium-mediated transformation. Cell, 2006,125(4):749-760. |
| [18] | NG D W K, ABEYSINGHE J K, KAMALI M . Regulating the regulators: The control of transcription factors in plant defense signaling. International Journal of Molecular Sciences, 2018,19(12):3737. |
| [19] | DIETZ K J, VOGEL M O, VIEHHAUSER A . AP2/EREBP transcription factors are part of gene regulatory networks and integrate metabolic, hormonal and environmental signals in stress acclimation and retrograde signalling. Protoplasma, 2010,245(1/4):3-14. |
| [20] | GAUTAM J K, NANDI A K . APD1, the unique member of Arabidopsis AP2 family influences systemic acquired resistance and ethylene-jasmonic acid signaling. Plant Physiology and Biochemistry, 2018,133:92-99. |
| [21] | JIN Y, PAN W, ZHENG X, CHENG X, LIU M, MA H, GE X . OsERF101, an ERF family transcription factor, regulates drought stress response in reproductive tissues. Plant Molecular Biology, 2018,98(1/2):51-65. |
| [22] | PANDEY S P, SOMSSICH I E . The role of WRKY transcription factors in plant immunity. Plant Physiology, 2009,150(4):1648-1655. |
| [23] | ZHENG Z, QAMAR S A, CHEN Z, MENGISTE T . Arabidopsis WRKY33 transcription factor is required for resistance to necrotrophic fungal pathogens. The Plant Journal, 2006,48(4):592-605. |
| [24] | CHUJO T, TAKAI R, AKIMOTO-TOMIYAMA C, ANDO S, MINAMI E, NAGAMURA Y, KAKU H, SHIBUYA N, YASUDA M, NAKASHITA H, UMEMURA K, OKADA A, OKADA K, NOJIRI H, YAMANE H . Involvement of the elicitor-induced gene OsWRKY53 in the expression of defense-related genes in rice. Biochimica et Biophysica Acta (BBA)-Gene Structure and Expression, 2007,1769(7/8):497-505. |
| [25] | DUBOS C, STRACKE R, GROTEWOLD E, WEISSHAAR B, MARTIN C, LEPINIEC L . MYB transcription factors in Arabidopsis. Trends in Plant Science, 2010,15(10):573-581. |
| [26] | LIU J, OSBOURN A, MA P . MYB transcription factors as regulators of phenylpropanoid metabolism in plants. Molecular Plant, 2015,8(5):689-708. |
| [27] | MA Q H, WANG C, ZHU H H . TaMYB4 cloned from wheat regulates lignin biosynthesis through negatively controlling the transcripts of both cinnamyl alcohol dehydrogenase and cinnamoyl-CoA reductase genes.Biochimie, 2011,93(7):1179-1186. |
| [28] | HUANG Y, LI T, XU Z S, WANG F, XIONG A S, HUANG Y, LI T, XU Z S, WANG F, XIONG A S . Six NAC transcription factors involved in response to TYLCV infection in resistant and susceptible tomato cultivars. Plant Physiology and Biochemistry, 2017,120:61-74. |
| [29] | YAN J, TONG T, LI X, CHEN Q, DAI M, NIU F, YANG M, DEYHOLOS M K, YANG B, JIANG Y Q . A novel NAC-type transcription factor, NAC87, from oilseed rape modulates reactive oxygen species accumulation and cell death. Plant and Cell Physiology, 2018,59(2):290-303. |
| [30] | WANG G, ZHANG S, MA X, WANG Y, KONG F, MENG Q . A stress-associated NAC transcription factor (SlNAC35) from tomato plays a positive role in biotic and abiotic stresses. Physiologia Plantarum, 2016,158(1):45-64. |
| [31] | VAN LOON L C, REP M, PIETERSE C M J . Significance of inducible defense-related proteins in infected plants. Annual Review of Phytopathology, 2006,44:135-162. |
| [32] | ANTONIW J F, RITTER C E, PIERPONT W S ,VAN LOON L C. Comparison of three pathogenesis-related proteins from plants of two cultivars of tobacco infected with TMV. Journal of General Virology, 1980,47(1):79-87. |
| [33] | VAN LOON L C . Regulation of changes in proteins and enzymes associated with active defence against virus infection//Active Defense Mechanisms in Plants, 1982,37:247-273. |
| [34] | LAGRIMINI L M, BURKHART W, MOYER M, ROTHSTEIN S . Molecular cloning of complementary DNA encoding the lignin- forming peroxidase from tobacco: Molecular analysis and tissue- specific expression. Proceedings of the National Academy of Sciences of the United States of America, 1987,84(21):7542-7546. |
| [35] | 李瑞博, 崔秀明, 刘玉忠, 吴志刚, 林淑芳, 申业, 黄璐琦 . 三七病程相关蛋白1基因的克隆与表达分析. 药学学报, 2014,49(1):124-130. |
| LI R B, CUI X M, LIU Y Z, WU Z G, LIN S F, SHEN Y, HUANG L Q . Cloning and expression analysis of pathogenesis-related protein 1 gene of Panax notoginseng. Acta Pharmaceutica Sinica, 2014,49(1):124-130. (in Chinese) | |
| [36] | MELCHERS L S, APOTHEKER-DE GROOT M,VAN DER KNAAP J A, PONSTEIN A S, SELA-BUURLAGE M B, BOL J F, CORNELISSEN B J, VAN DEN ELZEN P J, LINTHORST H J. , A new class of tobacco chitinases homologous to bacterial exo-chitinases displays antifungal activity. The Plant Journal, 1994,5(4):469-480. |
| [37] |
OKUSHIMA Y, KOIZUMI N, KUSANO T, SANO H . Secreted proteins of tobacco cultured BY2 cells: Identification of a new member of pathogenesis-related proteins. Plant Molecular Biology, 2000,42(3):479-488.
doi: 10.1023/A:1006393326985 |
| [1] | PEI YueHong,LI FengWei,LIU WeiNa,WEN YuXia,ZHU Xin,TIAN ShaoRui,FAN GuangJin,MA XiaoZhou,SUN XianChao. Characteristics of Cysteine Proteinase Gene Family in Nicotiana benthamiana and Its Function During TMV Infection [J]. Scientia Agricultura Sinica, 2022, 55(21): 4196-4210. |
| [2] | YuXia WEN,Jian ZHANG,Qin WANG,Jing WANG,YueHong PEI,ShaoRui TIAN,GuangJin FAN,XiaoZhou MA,XianChao SUN. Cloning, Expression and Anti-TMV Function Analysis of Nicotiana benthamiana NbMBF1c [J]. Scientia Agricultura Sinica, 2022, 55(18): 3543-3555. |
| [3] | MingYue GONG,XiaoTian DUAN,TingTing YU,Jie WANG,LiLi SHEN,Ying LI,MingHong LIU,YongLiang LI,HongKun LÜ,SongBai ZHANG,JinGuang YANG. Cloning of Hsc70-2 and Its Promoting Effect on Potato virus Y Infection in Nicotiana benthamiana [J]. Scientia Agricultura Sinica, 2020, 53(4): 771-781. |
| [4] | LIU HaiYang, WANG Wei, ZHANG RenFu, RAXIDA ·ABDURAHMAN, YAO Ju. Fungal Community Structure of Cotton-Field Soil Under Different Incidences of Cotton Verticillium Wilt [J]. Scientia Agricultura Sinica, 2019, 52(3): 455-465. |
| [5] | YUAN JunHu,DING YiJuan,YANG WenJing,YAN BaoQin,CHAI YaRu,MEI JiaQin,QIAN Wei. Identification of Genes Encoding Secretory Proteins Related to the Pathogenicity of Sclerotinia sclerotiorum Using TRV-HIGS [J]. Scientia Agricultura Sinica, 2019, 52(23): 4274-4284. |
| [6] | LIU RuiChi,CHENG YouPu,CHAI ALi,SHI YanXia,XIE XueWen, PATIGULI,LI BaoJu. Establishment and Application of a Triplex PCR Detection System for Vegetable Soil-Borne Pathogens [J]. Scientia Agricultura Sinica, 2019, 52(12): 2069-2078. |
| [7] | WU GenTu, CHEN GuangXiang, ZHANG JiaYuan, HU Qiao, MA MingGe, DOU YanXia, LI MingJun, QING Ling. Disease Resistance of Rice stripe virus NS3-Transgenic Nicotiana benthamiana [J]. Scientia Agricultura Sinica, 2019, 52(10): 1710-1720. |
|
||