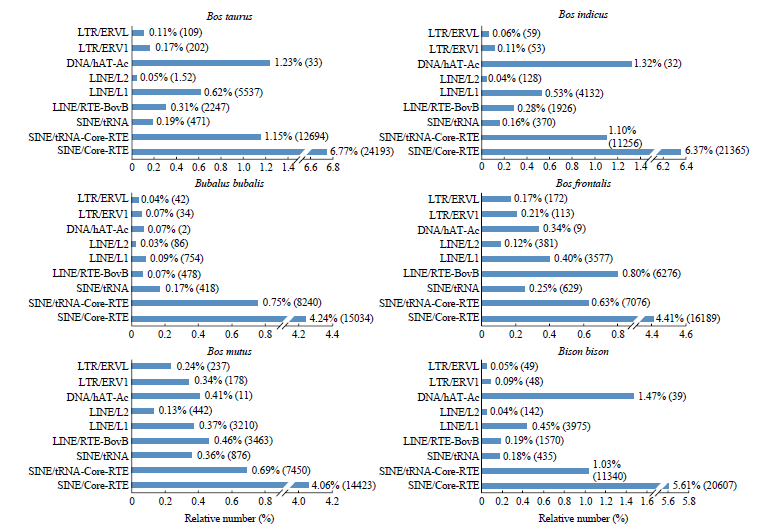
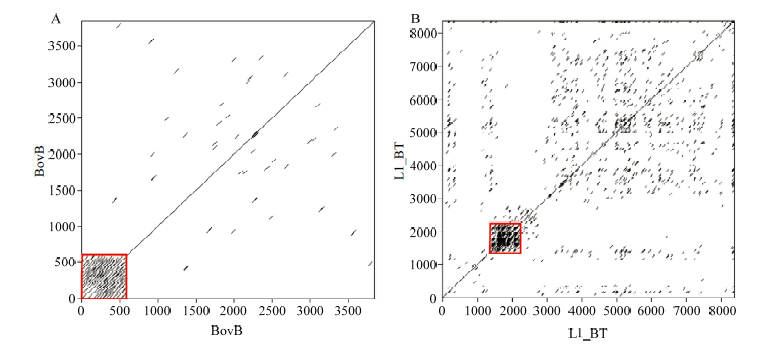
Scientia Agricultura Sinica ›› 2022, Vol. 55 ›› Issue (9): 1859-1867.doi: 10.3864/j.issn.0578-1752.2022.09.014
• ANIMAL SCIENCE·VETERINARY SCIENCE·RESOURCE INSECT • Previous Articles Next Articles
ZHANG Rui( ),ZHANG TianLiu,FAN TingTing,ZHU Bo,ZHANG LuPei,XU LingYang,GAO HuiJiang,LI JunYa,CHEN Yan,GAO Xue*()
),ZHANG TianLiu,FAN TingTing,ZHU Bo,ZHANG LuPei,XU LingYang,GAO HuiJiang,LI JunYa,CHEN Yan,GAO Xue*()
| [1] |
DE KONING A P J, GU W J, CASTOE T A, BATZER M A, POLLOCK D D. Repetitive elements may comprise over two-thirds of the human genome. PLoS Genetics, 2011, 7(12): e1002384. doi: 10.1371/journal.pgen.1002384.
doi: 10.1371/journal.pgen.1002384 |
| [2] |
SCHNABLE P S, WARE D, FULTON R S, STEIN J C, WEI F S, PASTERNAK S, LIANG C Z, ZHANG J W, FULTON L, GRAVES T A, et al. The B73 maize genome: Complexity, diversity, and dynamics. Science, 2009, 326(5956): 1112-1115. doi: 10.1126/science.1178534.
doi: 10.1126/science.1178534 |
| [3] |
HESLOP-HARRISON J S, SCHWARZACHER T. Organisation of the plant genome in chromosomes. The Plant Journal, 2011, 66(1): 18-33. doi: 10.1111/j.1365-313X.2011.04544.x.
doi: 10.1111/j.1365-313X.2011.04544.x. |
| [4] |
艾对元. 基因组中重复序列的意义. 生命的化学, 2008, 28(3): 343-345. doi: 10.3969/j.issn.1000-1336.2008.03.031.
doi: 10.3969/j.issn.1000-1336.2008.03.031 |
|
AI D Y. The meaning of repeat sequences. Chemistry of Life, 2008, 28(3): 343-345. doi: 10.3969/j.issn.1000-1336.2008.03.031. (in Chinese)
doi: 10.3969/j.issn.1000-1336.2008.03.031 |
|
| [5] |
AHMED M, LIANG P. Transposable elements are a significant contributor to tandem repeats in the human genome. Comparative and Functional Genomics, 2012, 2012: 947089. doi: 10.1155/2012/947089.
doi: 10.1155/2012/947089 |
| [6] |
JURKA J, GENTLES A J. Origin and diversification of minisatellites derived from human Alu sequences. Gene, 2006, 365: 21-26. doi: 10.1016/j.gene.2005.09.029.
doi: 10.1016/j.gene.2005.09.029 |
| [7] |
WONG L H, CHOO K H A. Evolutionary dynamics of transposable elements at the centromere. Trends in Genetics, 2004, 20(12): 611-616. doi: 10.1016/j.tig.2004.09.011.
doi: 10.1016/j.tig.2004.09.011 |
| [8] |
MACAS J, KOBLÍŽKOVÁ A, NAVRÁTILOVÁ A, NEUMANN P. Hypervariable 3' UTR region of plant LTR-retrotransposons as a source of novel satellite repeats. Gene, 2009, 448(2): 198-206. doi: 10.1016/j.gene.2009.06.014.
doi: 10.1016/j.gene.2009.06.014 |
| [9] |
SHARMA A, WOLFGRUBER T K, PRESTING G G. Tandem repeats derived from centromeric retrotransposons. BMC Genomics, 2013, 14: 142. doi: 10.1186/1471-2164-14-142.
doi: 10 |
| [10] |
CHENG Z J, MURATA M. A centromeric tandem repeat family originating from a part of Ty3/Gypsy-retroelement in wheat and its relatives. Genetics, 2003, 164(2): 665-672. doi: 10.1093/genetics/164.2.665.
doi: 10.1093/genetics/164.2.665 |
| [11] |
MILLER W J, NAGEL A, BACHMANN J, BACHMANN L.Evolutionary dynamics of the SGM transposon family in the Drosophila obscura species group. Molecular Biology and Evolution, 2000, 17(11): 1597-1609. doi: 10.1093/oxfordjournals.molbev.a026259.
doi: 10.1093/oxfordjournals.molbev.a026259 |
| [12] |
PONTECORVO G, DE FELICE B, CARFAGNA M. A novel repeated sequence DNA originated from a Tc1-like transposon in water green frog Rana esculenta. Gene, 2000, 261(2): 205-210. doi: 10.1016/S0378-1119(00)00539-4.
doi: 10.1016/S0378-1119(00)00539-4 |
| [13] |
KAPITONOV V V, HOLMQUIST G P, JURKA J. L1 repeat is a basic unit of heterochromatin satellites in cetaceans. Molecular Biology and Evolution, 1998, 15(5): 611-612. doi: 10.1093/oxfordjournals.molbev.a025963.
doi: 10.1093/oxfordjournals.molbev.a025963 |
| [14] |
GAFFNEY P M, PIERCE J C, MACKINLEY A G, TITCHEN D A, GLENN W K. Pearl, a novel family of putative transposable elements in bivalve mollusks. Journal of Molecular Evolution, 2003, 56(3): 308-316. doi: 10.1007/s00239-002-2402-5.
doi: 10.1007/s00239-002-2402-5 |
| [15] |
BOVINE G S, ANALYSIS C, ELSIK C G, GIBBS R A, MUZUNY D M, WEINSTOCK G M, AELSON D L, EICHLER E E, ELNITSKI L, GUIGO R, et al. The genome sequence of taurine cattle: A window to ruminant biology and evolution. Science, 2009, 324(5926): 522-8.
doi: 10.1126/science.1169588 |
| [16] |
WANG K, WANG L Z, LENSTRA J A, JIAN J B, YANG Y Z, HU Q J, LAI D Y, QIU Q, MA T, DU Z, ABBOTT R, LIU J Q. The genome sequence of the wisent (Bison bonasus). GigaScience, 2017, 6(4): gix016. doi: 10.1093/gigascience/gix016.
doi: 10.1093/gigascience/gix016 |
| [17] |
WANG M S, ZENG Y, WANG X, NIE W H, WANG J H, SU W T, OTECKO N O, XIONG Z J, WANG S, QU K X, YAN S Q, YANG M M, WANG W, DONG Y, WU D D, ZHANG Y P. Draft genome of the gayal, Bos frontalis. GigaScience, 2017, 6(11): gix094. doi: 10.1093/gigascience/gix094.
doi: 10.1093/gigascience/gix094 |
| [18] |
GLANZMANN B, MÖLLER M, LE ROEX N, TROMP G, HOAL E G, VAN HELDEN P D. The complete genome sequence of the African buffalo (Syncerus caffer). BMC Genomics, 2016, 17(1): 1001. doi: 10.1186/s12864-016-3364-0.
doi: 10.1186/s12864-016-3364-0 |
| [19] |
BENSON G. Tandem repeats finder: A program to analyze DNA sequences. Nucleic Acids Research, 1999, 27(2): 573-580. doi: 10.1093/nar/27.2.573.
doi: 10.1093/nar/27.2.573 |
| [20] |
MELTERS D P, BRADNAM K R, YOUNG H A, TELIS N, MAY M R, RUBY J G, SEBRA R, PELUSO P, EID J, RANK D, GARCIA J F, DERISI J L, SMITH T, TOBIAS C, ROSS-IBARRA J, KORF I, CHAN S W L. Comparative analysis of tandem repeats from hundreds of species reveals unique insights into centromere evolution. Genome Biology, 2013, 14(1): R10. doi: 10.1186/gb-2013-14-1-r10.
doi: 10 |
| [21] |
AMES D, MURPHY N, HELENTJARIS T, SUN N N, CHANDLER V. Comparative analyses of human single- and multilocus tandem repeats. Genetics, 2008, 179(3): 1693-1704. doi: 10.1534/genetics.108.087882.
doi: 10.1534/genetics.108.087882 |
| [22] |
MEŠTROVIĆ N, MRAVINAC B, PAVLEK M, VOJVODA-ZELJKO T, ŠATOVIĆ E, PLOHL M. Structural and functional liaisons between transposable elements and satellite DNAs. Chromosome Research, 2015, 23(3): 583-596. doi: 10.1007/s10577-015-9483-7.
doi: 10.1007/s10577-015-9483-7 |
| [23] |
VONDRAK T, ROBLEDILLO L Á, NOVÁK P, KOBLÍŽKOVÁ A, NEUMANN P, MACAS J. Characterization of repeat arrays in ultra-long nanopore reads reveals frequent origin of satellite DNA from retrotransposon-derived tandem repeats. The Plant Journal: for Cell and Molecular Biology, 2020, 101(2): 484-500. doi: 10.1111/tpj.14546.
doi: 10.1111/tpj.14546 |
| [24] |
LÓPEZ-FLORES I, GARRIDO-RAMOS M A. The repetitive DNA content of eukaryotic genomes. Genome Dynamics, 2012, 7: 1-28. doi: 10.1159/000337118.
doi: 10.1159/000337118 |
| [25] |
CHENG Z K, DONG F G, LANGDON T, OUYANG S, BUELL C R, GU M H, BLATTNER F R, JIANG J M. Functional rice centromeres are marked by a satellite repeat and a centromere-specific retrotransposon. The Plant Cell, 2002, 14(8): 1691-1704. doi: 10.1105/tpc.003079.
doi: 10.1105/tpc.003079 |
| [26] |
ZHONG C X, MARSHALL J B, TOPP C, MROCZEK R, KATO A, NAGAKI K, BIRCHLER J A, JIANG J M, DAWE R K. Centromeric retroelements and satellites interact with maize kinetochore protein CENH3. The Plant Cell, 2002, 14(11): 2825-2836. doi: 10.1105/tpc.006106.
doi: 10.1105/tpc.006106 |
| [27] |
LIU Z, YUE W, LI D Y, WANG R R C, KONG X Y, LU K, WANG G X, DONG Y S, JIN W W, ZHANG X Y. Structure and dynamics of retrotransposons at wheat centromeres and pericentromeres. Chromosoma, 2008, 117(5): 445-456. doi: 10.1007/s00412-008-0161-9.
doi: 10.1007/s00412-008-0161-9 |
| [28] |
LANGDON T, SEAGO C, JONES R N, OUGHAM H, THOMAS H, FORSTER J W, JENKINS G. De novo evolution of satellite DNA on the rye B chromosome. Genetics, 2000, 154(2): 869-884. doi: 10.1093/genetics/154.2.869.
doi: 10.1093/genetics/154.2.869 |
| [29] |
GONG Z Y, WU Y F, KOBLÍŽKOVÁ A, TORRES G A, WANG K, IOVENE M, NEUMANN P, ZHANG W L, NOVÁK P, BUELL C R, MACAS J, JIANG J M. Repeatless and repeat-based centromeres in potato: implications for centromere evolution. The Plant Cell, 2012, 24(9): 3559-3574. doi: 10.1105/tpc.112.100511.
doi: 10.1105/tpc.112.100511 |
| [30] |
HIKOSAKA A, KAWAHARA A. Lineage-specific tandem repeats riding on a transposable element of MITE in Xenopus evolution: A new mechanism for creating simple sequence repeats. Journal of Molecular Evolution, 2004, 59(6): 738-746. doi: 10.1007/s00239-004-2664-1.
doi: 10.1007/s00239-004-2664-1 |
| [31] |
PLOHL M, MEŠTROVIĆ N, MRAVINAC B. Satellite DNA evolution. Genome Dynamics, 2012, 7:126-152. doi: 10.1159/000337122.
doi: 10.1159/000337122 |
| [32] |
MCGURK M P, BARBASH D A. Double insertion of transposable elements provides a substrate for the evolution of satellite DNA. Genome Research, 2018, 28(5): 714-725. doi: 10.1101/gr.231472.117.
doi: 10.1101/gr.231472.117 |
| [33] |
KAPITONOV V V, JURKA J. Molecular paleontology of transposable elements from Arabidopsis thaliana. Genetica, 1999, 107(1/2/3): 27-37.
doi: 10.1023/A:1004030922447 |
| [34] |
SMIT A F A, RIGGS A D. MIRs are classic, tRNA-derived SINEs that amplified before the mammalian radiation. Nucleic Acids Research, 1995, 23(1): 98-102. doi: 10.1093/nar/23.1.98.
doi: 10.1093/nar/23.1.98 |
| [35] | DAMIANI G, FLORIO S, PANELLI S, CAPELLI E, CUCCIA M. The Bov-A2 retroelement played a crucial role in the evolution of ruminants. Rivista Di Biologia, 2008, 101(3): 375-404. |
| [36] |
YANG H P, BARBASH D A. Abundant and species-specific DINE-1 transposable elements in 12 Drosophila genomes. Genome Biology, 2008, 9(2): R39. doi: 10.1186/gb-2008-9-2-r39.
doi: 10 |
| [37] |
THOMAS J, VADNAGARA K, PRITHAM E J. DINE-1, the highest copy number repeats in Drosophila melanogaster are non-autonomous endonuclease-encoding rolling-circle transposable elements (Helentrons). Mobile DNA, 2014, 5: 18. doi: 10.1186/1759-8753-5-18.
doi: 10.1186/1759-8753-5-18 |
| [38] |
LUCHETTI A. terMITEs: miniature inverted-repeat transposable elements (MITEs) in The Termite Genome (Blattodea: Termitoidae). Molecular Genetics and Genomics: MGG, 2015, 290(4): 1499-1509. doi: 10.1007/s00438-015-1010-1.
doi: 10.1007/s00438-015-1010-1 |
| [39] |
NOMA K. Tnat1 and Tnat 2 from Arabidopsis thaliana: novel transposable elements with tandem repeat sequences. DNA Research, 2000, 7(1): 1-7. doi: 10.1093/dnares/7.1.1.
doi: 10.1093/dnares/7.1.1 |
| [40] |
SCALVENZI T, POLLET N. Insights on genome size evolution from a miniature inverted repeat transposon driving a satellite DNA. Molecular Phylogenetics and Evolution, 2014, 81: 1-9. doi: 10.1016/j.ympev.2014.08.014.
doi: 10.1016/j.ympev.2014.08.014 |
| [41] |
DIAS G B, SVARTMAN M, DELPRAT A, RUIZ A, KUHN G C S. Tetris is a foldback transposon that provided the building blocks for an emerging satellite DNA of Drosophila virilis. Genome Biology and Evolution, 2014, 6(6): 1302-1313. doi: 10.1093/gbe/evu108.
doi: 10.1093/gbe/evu108 |
| [42] |
MARTÍNEZ-IZQUIERDO J A, GARCÍA-MARTÍNEZ J, VICIENT C M. What makes Grande1 retrotransposon different? Genetica, 1997, 100(1/2/3): 15-28.
doi: 10.1023/A:1018332218319 |
| [1] | MIAO JiaHao, CUI PengFei, YAN Cheng, WANG CongCong, WANG Yan, CHEN Yuan, CHEN Peng, SHI JianZhong, DENG GuoHua, CHEN HuaLan. Genetic and Biological Characterization of Two H8N4 Subtype Avian Influenza Viruses Isolated from Duck [J]. Scientia Agricultura Sinica, 2026, 59(7): 1576-1586. |
| [2] | ZHANG TianYu, LIU LiJun, YANG Gang, WU JunYan, PU YuanYuan, MA Li, WANG WangTian, LU XiaoMing, MA YuanQiang, SUN WanCang. Genetic Gain Analysis of Agronomic Traits of Brassica rapa L. in Northern China from 2006 to 2022 [J]. Scientia Agricultura Sinica, 2025, 58(11): 2081-2095. |
| [3] | ZHU QingYing, HE GengYi, SUN Ping. Research on the Spatio-Temporal Evolution and Differentiated Improvement Path of Low-Carbon Utilization Efficiency of Cultivated Land Under the Background of New and Old Kinetic Energy Conversion——A Case Study of Guangdong Province [J]. Scientia Agricultura Sinica, 2025, 58(11): 2206-2224. |
| [4] | WEI NaiCui, TAO JinBo, YUAN MingYang, ZHANG Yu, KAI MengXiang, QIAO Ling, WU BangBang, HAO YuQiong, ZHENG XingWei, WANG JuanLing, ZHAO JiaJia, ZHENG Jun. Seedling Characterization and Genetic Analysis of Low Phosphorus Tolerance in Shanxi Varieties [J]. Scientia Agricultura Sinica, 2024, 57(5): 831-845. |
| [5] | LI BOTIAN, LI CHUNQI, LIU GUOPING, XIE JUN, ZENG PAN, ZHAO RUNZE, LI TONG, PEI JIE, GUO LIWEI, WU RUI, TAN LEI. Molecular Epidemic Characteristics and Genetic Variation Analysis of Porcine Circovirus 3 in Some Pig Farms in Central China from 2020 to 2022 [J]. Scientia Agricultura Sinica, 2024, 57(3): 613-626. |
| [6] | LI XinLei, SUN JiuYing, YANG Cheng, CHENG Ning, WANG KaiYue, WANG HuanHuan, CHENG XueJiao, ZHAO Jian, SUN YingFeng. Isolation, Identification and Genetic Variation of a Three-Lineage Strain Recombined Porcine Reproductive and Respiratory Syndrome Virus [J]. Scientia Agricultura Sinica, 2024, 57(24): 4978-4989. |
| [7] | LI Pei, HE ZhiLin, TAN YueXia, ZHAO WanTong, FENG JinYing, CHEN GuiHu, YAN Chi, WANG ZiHao, HUANG Ping, JIANG Dong. Genetic Diversity Analysis of Mandarin and Excellent Germplasm Screening Based on Whole-Genome Resequencing Data and Phenotypic Traits [J]. Scientia Agricultura Sinica, 2024, 57(23): 4761-4773. |
| [8] | REN CaiXia, LIU Lin, LIU ShengXue, BU Fang DI, XIANG BenChun, ZHENG YinYing, CUI BaiMing. Complete Genome Sequence Analysis and Infectious Clone Construction of Mume Virus A Peach Isolate pp in Xinjiang [J]. Scientia Agricultura Sinica, 2024, 57(19): 3810-3822. |
| [9] | YAN LiuHui, ZHONG Qi, MA ZengFeng, WEI MinYi, LIU Chi, QIN YuanYuan, ZHOU XiaoLong, HUANG DaHui, LU YingPing, QIN Gang, ZHANG YueXiong. Identification and Evolutionary Analysis of the Early Heading Gene OsEHD8 in Common Wild Rice (Oryza rufipogon Giff.) [J]. Scientia Agricultura Sinica, 2024, 57(14): 2703-2716. |
| [10] | MENG QingLei, YIN YuXiang, WANG YuHao. Spatial-Temporal Evolution, Decoupling Effect and Performance Evaluation of China’s Agricultural Carbon Emissions [J]. Scientia Agricultura Sinica, 2023, 56(20): 4049-4066. |
| [11] | WANG CaiXiang,YUAN WenMin,LIU JuanJuan,XIE XiaoYu,MA Qi,JU JiSheng,CHEN Da,WANG Ning,FENG KeYun,SU JunJi. Comprehensive Evaluation and Breeding Evolution of Early Maturing Upland Cotton Varieties in the Northwest Inland of China [J]. Scientia Agricultura Sinica, 2023, 56(1): 1-16. |
| [12] | HUA ChunLin,ZHANG JiuHong,JIN ShuQin. Analysis to Evolution Characteristics of Policies for Controlling Agricultural Non-Point Source Pollution in China: Based on Text Quantification [J]. Scientia Agricultura Sinica, 2022, 55(7): 1385-1398. |
| [13] | WANG YanWen,WANG MengJing,ZHANG Hong,GAO XinXin,GUO Jing,LI XuYong. Evolution of Human H9N2 Avian Influenza Virus in China from 1998 to 2021 [J]. Scientia Agricultura Sinica, 2022, 55(20): 4075-4090. |
| [14] | YANG Cheng,GONG GuiZhi,PENG ZhuChun,CHANG ZhenZhen,YI Xuan,HONG QiBin. Genetic Relationship Among Citrus and Its Relatives as Revealed by cpInDel and cpSSR Marker [J]. Scientia Agricultura Sinica, 2022, 55(16): 3210-3223. |
| [15] | XIE Bin,AN XiuHong,CHEN YanHui,CHENG CunGang,KANG GuoDong,ZHOU JiangTao,ZHAO DeYing,LI Zhuang,ZHANG YanZhen,YANG An. Response and Adaptability Evaluation of Different Apple Rootstocks to Continuous Phosphorus Deficiency [J]. Scientia Agricultura Sinica, 2022, 55(13): 2598-2612. |
|
||