






中国农业科学 ›› 2024, Vol. 57 ›› Issue (3): 429-441.doi: 10.3864/j.issn.0578-1752.2024.03.001
张必东1,2( ), 林泓1, 朱思颖1, 李忠成1, 庄慧1, 李云峰1()
), 林泓1, 朱思颖1, 李忠成1, 庄慧1, 李云峰1()
收稿日期:2023-07-05
接受日期:2023-08-28
出版日期:2024-02-01
发布日期:2024-02-05
通信作者:
联系方式:
张必东,E-mail:vacom30@163.com。
基金资助:
ZHANG BiDong1,2(), LIN Hong1, ZHU SiYing1, LI ZhongCheng1, ZHUANG Hui1, LI YunFeng1()
Received:2023-07-05
Accepted:2023-08-28
Published:2024-02-01
Online:2024-02-05
摘要:
【目的】水稻是世界性的粮食作物,其籽粒形态直接影响水稻的最终产量和营养品质,进而影响其经济价值。此外,水稻花发育与籽粒形态又具有复杂的相关性,探究新的水稻花发育调控基因及其分子调控机制可以为培育籽粒更大、更饱满的水稻品种奠定基础。【方法】在利用甲基磺酸乙酯(ethyl methyl sulfonate,EMS)诱变西大1B(籼稻保持系)得到的突变体库中鉴定到一个颖壳和浆片发育异常且矮化的突变体abnormal hull 1(ah1);观察并统计野生型和突变体的农艺性状;选取各个时期的小穗,对突变体的组织学、形态学变化进行分析;采用ah1和籼稻温敏不育系56S构建F2分离群体,并将其用于遗传分析和基因定位;提取野生型和突变体的幼穗RNA并将其反转录为cDNA,对花发育调控基因和ABA合成关键基因的表达量进行RT-qPCR分析。【结果】农艺性状分析表明,突变体ah1各节间的大幅缩短直接导致其矮化;同时,突变体的小穗畸形严重,结实率极低。组织学和形态学分析发现,ah1小穗主要表现为内外稃、浆片和雄蕊等花器官发生了不同程度的退化,部分严重的小穗出现了花器官特征和花分生组织确定性的改变,且伴随大面积的白化,根据其退化程度的不同可分为一般和严重2种类型。遗传分析发现分离群体中野生型和突变体的比值为3﹕1,表明ah1突变性状受1个隐性基因控制。AH1定位于第1染色体上的分子标记RM6716和RM128之间,物理距离近8 Mb,对突变体进行重测序分析后发现该区间内的LOC_Os01g53450和LOC_Os01g51860在野生型和突变体之间出现了变异,因此,将这两个基因暂定为候选基因。RT-qPCR分析表明,在突变体幼穗早期发育过程中,各花器官发育调控基因的表达量发生了显著的变化;同时,ABA合成关键基因OsNCED1/OsNCED2/OsNCED3/OsNCED4/OsNCED5的表达严重受阻。【结论】AH1对于维持水稻内外稃等花器官的形态建成起到至关重要的作用,将LOC_Os01g53450和LOC_Os01g51860暂定为候选基因。
张必东, 林泓, 朱思颖, 李忠成, 庄慧, 李云峰. 水稻颖壳异常突变体ah1的鉴定与候选基因分析[J]. 中国农业科学, 2024, 57(3): 429-441.
ZHANG BiDong, LIN Hong, ZHU SiYing, LI ZhongCheng, ZHUANG Hui, LI YunFeng. Identification and Candidate Gene Analysis of the ABNORMAL HULL 1 (ah1) Mutant in Rice (Oryza sativa L.)[J]. Scientia Agricultura Sinica, 2024, 57(3): 429-441.
表1
引物序列"
| 引物 Primer | 正向序列 Forward sequence (5′-3′) | 反向序列 Reverse sequence (5′-3′) | 用途 Purpose |
|---|---|---|---|
| RM6716 | CCTATGCGATCTCCTATTTGG | CGAGTCAGACACGTACTACTGC | 定位 Mapping |
| RM128 | TGATTTCTTGGAAGCGAAGAGTGAGG | CCTCCTTGTGCTCAGCCATGC | |
| RM1268 | GCTGTCACTGACCGAGCGTAGG | TCGAGAGATCCAATCCAGTTTGC | |
| ACTIN | GACCCAGATCATGTTTGAGACCT | CAGTGTGGCTGACACCATCAC | RT-qPCR |
| OsMADS14 | CTCATCATCCTCCTTCAT | GCTACATCCTCTATCCTT | |
| OsMADS15 | GCTGAATCCGAGAGTGAG | TGAGGTGTTTGTGACATTTG | |
| OsMADS18 | GGTGATGAATATGGAATT | CTTATATGCTTCAGAGAAT | |
| OsMADS2 | ATGATTGAGGAGGCACTT | TCGTCTTCCAGCATCTTAT | |
| OsMADS4 | CCAATCTGCGGGACAAGA | AGCCAAATTGGCAGTGCTC | |
| OsMADS16 | TTCGCCACAGGAAGTATCA | CCTCGTAGGAGTGCTTCA | |
| OsMADS3 | CGATCATATGGTTAATAACCCA | AGACGAGTTGCCAAAGTTCGA | |
| OsMADS58 | ATATCACCACCATGAACCA | TTCTTCCGTGCTCTAATCT | |
| OsMADS13 | AGAGCCAGGAAGAATGAA | ATGTTGTCGTTCTGAAGC | |
| OsMADS1 | GCTGCAACTACAACTCACAGG | TGATGGTGAGCATGAGGGTG | |
| OsMADS5 | ACTGAACTAAGCAATTACCAAGA | CAAGTCCTCGCCAAGAAG | |
| OsMADS7 | ATCTTGATTCATTAGGCATAA | TTGTCCTTGTAGTTCTGA | |
| OsMADS8 | GCACATTAGATCCACAAG | TTCTTCTGAGGCACTTAT | |
| OsMADS34 | AGGAATATGTGAACTTGAA | TTCTGACTACTTGACTCT | |
| DL | CCCATCTGCTTACAACCGCTT | GTTGGAGGTGGAAACCGTCG | |
| OsMADS6 | TGATGATGGAACAAGTGGAG | TTGTGCTTGAGTTGCCTAT | |
| OsMADS32 | GGAGTTTGTCGAGAGGTACGA | CTGAACGTCAAGCGTCCATC | |
| SL1 | AAGCATCCAGGGTTACTAA | GCAGAACCTACACTCGTA | |
| FON1 | TGCGTGAAGGAGGACAACATCATC | AGCAGCAGGTTCGTCTCCCTGTT | |
| OsNCED1 | AAGCAGACCGAGCAAGAG | ATTCGTCGTAGCAATAACAGTTC | |
| OsNCED2 | AGCCAATGTTATGACTCT | TCTGTAGATTACGGTGAG | |
| OsNCED3 | AGGATATGCTCACATACAG | AGAATCTCACCGAATTGG | |
| OsNCED4 | ACGATGATCCACGACTTC | ATCTCCTGGAGCTTGAAC | |
| OsNCED5 | ATCAAGAAGCCATACCTGAA | CTGCTCAAGCTCGATCTC | |
| LOC_Os01g51860 | TGAAGGAATCTGCTGAAGT | AGTATCTGAGGACCACAATT | |
| LOC_Os01g53450 | ACGGAATGGCACAGTGGGAC | GGTCCCATGCCGTACTCTTGAG |

图1
野生型(WT)和突变体ah1的小穗表型分析 A1:野生型小穗;A2:去除内外稃后的野生型小穗;A3:Sp6期的野生型花原基;A4:Sp8期的野生型花原基;A5:野生型小穗的横切面;B1:ah1的小穗;B2:去除内外稃后的ah1的小穗;B3:Sp6期一般表型的突变体的花原基;B4:Sp8期一般表型的突变体的花原基;B5:一般表型的ah1的小穗横切面;C1和C2:突变表型严重的完整小穗及去除内外稃后的小穗;C3:Sp6期严重表型的突变体的花原基;C4:Sp8期严重表型的突变体的花原基;C5:严重表型的ah1的小穗横切面;D1和D2:内稃伸长、外稃退化、浆片异常发育的ah1小穗(类型Ⅰ);D3:退化的内稃,D2红框内的放大图;D4:D2小穗中异常发育的浆片;D5:D1小穗退化内稃的横切图;D6:D1小穗异常浆片的横切图;E1和E2:雄蕊雌蕊化、内外稃退化、浆片伸长的ah1小穗(类型Ⅱ);E3:退化的外稃,E2红框内的放大图;E4:E2小穗中伸长的浆片;E5:ah1中的双花小穗(类型Ⅲ)。le:外稃;pa:内稃;lo:浆片;st:雄蕊;pi:雌蕊;sl:护颖;rg:副护颖;mrp:内稃边缘;dl:退化的外稃;ele:伸长的外稃;alo:异常的浆片;elo:伸长的浆片;ex-floret:额外的小花;*:雄蕊。标尺:A1和A2均为5 000 µm;B1、B2、C1、C2、D1、E1和E5均为1 000 µm;A4、A5、B4、B5、C4、C5、D4、D5和D6均为500 µm;D2、E2均为2 000 µm;A3、B3、C3、D3、E3和E4均为100 µm"

图2
野生型(WT)和ah1突变体的株型及农艺性状 A:野生型与突变体的成熟期植株;B和C:野生型与突变体的稻穗(Ⅰ、Ⅱ、Ⅲ和Ⅳ分别代表倒一节间、倒二节间、倒三节间和倒四节间),标尺:5 cm;D:株高统计;E:节间数统计;F:结实率统计;G:千粒重统计。**:t检验中P<0.01,差异极显著。下同"
表2
野生型和ah1突变体分离的卡方测验"
| 父本 Male parent | 母本 Female parent | 野生型 Wild-type | 突变型 Mutant type | 卡方值 Chi-square value (χ20.05,1=3.84) |
|---|---|---|---|---|
| ah1 | 56S | 303 | 97 | 0.083 |

图3
AH1候选基因的定位和表达分析 A:AH1在第一染色体上的初步定位;B、C:LOC_Os01g53450和LOC_Os01g51860在小于3 mm的幼穗中的相对表达量;D、E:LOC_Os01g53450和LOC_Os01g51860在小于1 cm的幼穗中的相对表达量。F:野生型和突变体中的ABA含量"
表3
突变基因"
| 序号 Number | 基因名称 Gene name | 注释 Annotation | 位置 Position (Mb) |
|---|---|---|---|
| 1 | LOC_Os01g51860 | 紫黄素脱环氧化酶,推测,表达 Violaxanthin de-epoxidase, putative, expressed | 28.44 |
| 2 | LOC_Os01g53450 | 转氨酶,Ⅰ类和Ⅱ类,结构域蛋白,表达 Aminotransferase, classesⅠandⅡ, domain containing protein, expressed | 29.29 |
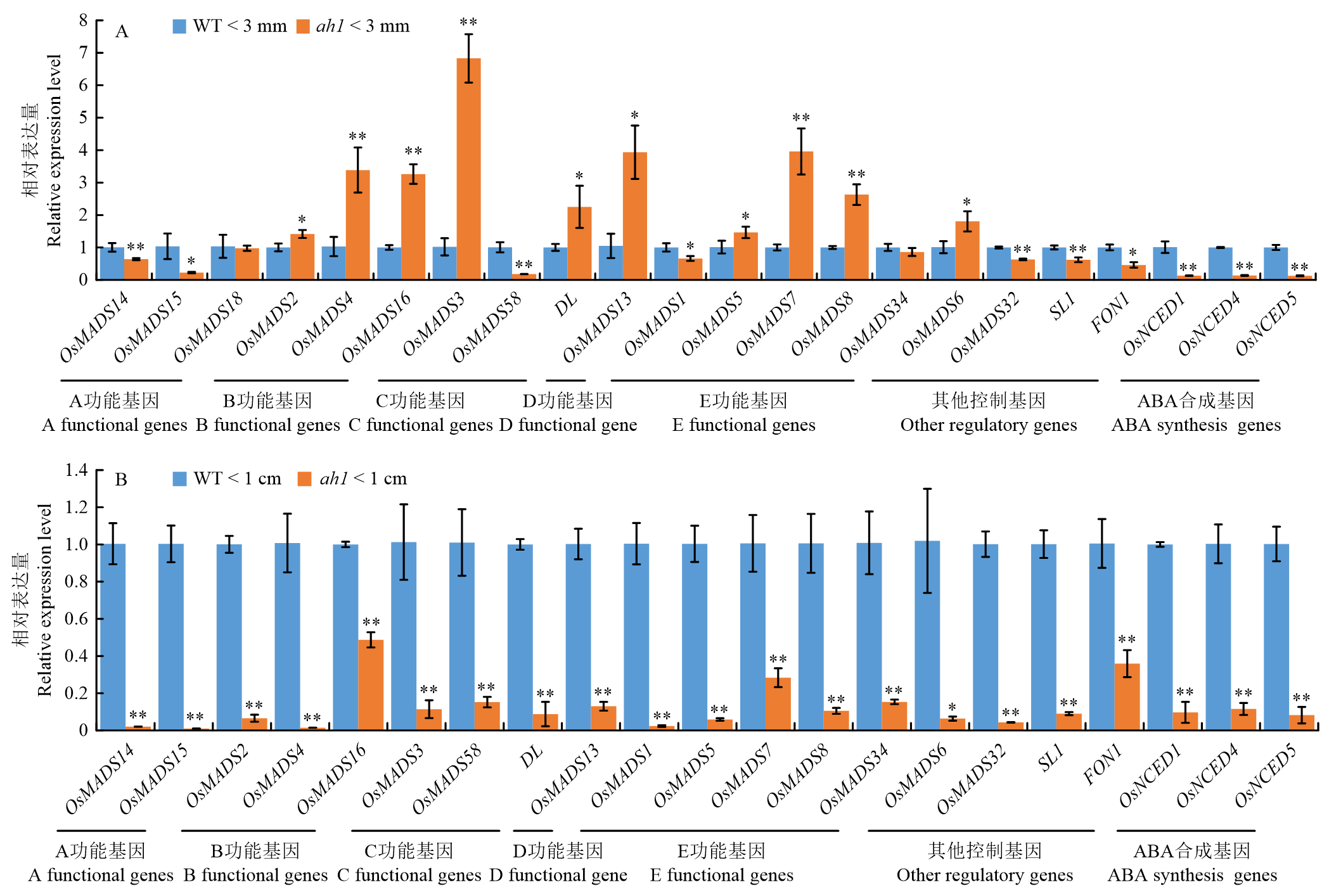
图4
水稻花发育调控基因和ABA合成基因在野生型和ah1突变体中的相对表达量 A:小于3 mm的幼穗中花器官发育调控基因和ABA合成基因的相对表达量(OsNCED2/OsNCED3几乎无表达,数据未展示);B:小于1 cm的幼穗中花器官发育调控基因和ABA合成基因的相对表达量(OsMADS18和OsNCED2/OsNCED3几乎无表达,数据未展示)。*:t检验中P<0.05差异显著"
| [1] |
doi: 10.1038/353031a0 |
| [2] |
doi: 10.1093/pcp/pcf010 |
| [3] |
pmid: 11832239 |
| [4] |
doi: 10.1093/aob/mcm143 pmid: 17670752 |
| [5] |
pmid: 17616563 |
| [6] |
pmid: 12815719 |
| [7] |
doi: 10.1007/s00497-002-0151-7 |
| [8] |
doi: 10.1371/journal.pgen.1000818 |
| [9] |
doi: 10.1104/pp.104.045039 |
| [10] |
doi: 10.1105/tpc.112.097105 |
| [11] |
doi: 10.1242/dev.00294 |
| [12] |
doi: 10.1023/A:1025401611354 |
| [13] |
doi: 10.1007/s00425-003-1066-8 |
| [14] |
doi: 10.1534/genetics.107.071746 |
| [15] |
doi: 10.1093/pcp/pcn050 |
| [16] |
doi: 10.1093/genetics/165.4.2301 |
| [17] |
doi: 10.1023/a:1006051911291 pmid: 9869408 |
| [18] |
doi: 10.1105/tpc.105.037200 |
| [19] |
doi: 10.1105/tpc.111.087007 pmid: 21810995 |
| [20] |
doi: 10.1093/mp/sst003 |
| [21] |
doi: 10.1105/tpc.018044 |
| [22] |
doi: 10.1111/tpj.2007.52.issue-4 |
| [23] |
|
| [24] |
|
| [25] |
doi: 10.1111/tpj.2005.43.issue-6 |
| [26] |
|
| [27] |
doi: 10.1007/s00425-005-0141-8 |
| [28] |
doi: 10.1007/s11103-005-2161-y |
| [29] |
doi: 10.1016/S1016-8478(23)13260-2 |
| [30] |
doi: 10.1111/j.1365-313X.2009.04101.x pmid: 20003164 |
| [31] |
doi: 10.1104/pp.110.156711 |
| [32] |
doi: 10.1007/s11103-012-9958-2 |
| [33] |
doi: 10.1105/tpc.109.068742 |
| [34] |
doi: 10.1104/pp.112.200980 pmid: 22891238 |
| [35] |
doi: 10.1016/j.ydbio.2005.03.016 |
| [36] |
doi: 10.1111/j.1365-313X.2009.03913.x pmid: 19453444 |
| [37] |
|
| [38] |
doi: 10.1016/j.jplph.2014.06.022 |
| [39] |
pmid: 16511358 |
| [40] |
doi: 10.1007/s00425-021-03802-1 pmid: 34842977 |
| [41] |
doi: 10.1016/j.pestbp.2015.05.006 |
| [42] |
pmid: 16299174 |
| [43] |
doi: 10.1111/j.1365-313X.2011.04871.x pmid: 22126457 |
| [1] | 彭廷燊, 陆久焱, 吴美林, 严雨欣, 刘宏周, 南文斌, 秦小健, 李明, 龚俊义, 梁永书. 多年生水稻黄糯2号和长白7号产量相关性状的QTL分析[J]. 中国农业科学, 2026, 59(7): 1361-1379. |
| [2] | 叶美金, 吴雷, Lohani Md Nahibuzzaman, 尹丽, 胡欣荣, 刘亚西, 蒋云峰, 陈国跃, 蒲至恩, 李阳, 李婷, 邹亚亚, 吴佳怡, 马建. 基于GWAS的中国地方小麦成熟胚大小位点的鉴定及其遗传效应解析[J]. 中国农业科学, 2026, 59(6): 1157-1171. |
| [3] | 陈敏, 焦紫岚, 乔承彬, 许昊, 张碧, 马东花, 孔维儒, 王敬文, 宋佳伟, 罗成科, 李培富, 田蕾. 不同亚种水稻种质资源对盐胁迫的形态生理响应及适应策略[J]. 中国农业科学, 2026, 59(4): 705-722. |
| [4] | 郭富城, 唐海江, 郝馨怡, 马国林, 杨九菊, 黄霖锋, 田蕾, 王彬, 罗成科. 不同灌溉方式对宁夏盐渍化土壤水盐运移、水稻产量及水分利用效率的影响[J]. 中国农业科学, 2026, 59(4): 750-764. |
| [5] | 张卫建, 严圣吉, 尚子吟, 唐志伟, 吴柳格, 李佳锐, 陈浩天, 邓艾兴, 张俊, 张鑫, 郑成岩, 宋振伟. 稻田甲烷不完全源于水稻种植的人为贡献[J]. 中国农业科学, 2026, 59(4): 824-833. |
| [6] | 朱姝, 国志鹏, 孙颖. 水稻雷帕霉素靶蛋白OsTOR调控根伸长的功能研究[J]. 中国农业科学, 2026, 59(3): 475-485. |
| [7] | 王勇胜, 牛丽, 王长杰, 马立花, 廉潇潇, 孟亚雄, 马小乐, 姚立蓉, 张宏, 杨轲, 李葆春, 王化俊, 司二静, 汪军成. 冬小麦千粒重的全基因组关联分析及候选基因预测[J]. 中国农业科学, 2026, 59(3): 499-514. |
| [8] | 吕文彦, 程海涛, 马兆惠, 田淑华. 育种新时代水稻杂交育种技术与策略探讨[J]. 中国农业科学, 2026, 59(2): 233-238. |
| [9] | 廖婷璐, 石亚飞, 肖东浩, 舍杨梦斐, 郭富城, 杨九菊, 唐海江, 罗成科. 外源硝普钠对碱胁迫下水稻幼苗糖代谢的影响[J]. 中国农业科学, 2026, 59(2): 265-277. |
| [10] | 刘天生, 刘耕源, 赵安琪, 杨旭, 蔡明雪, 杨艾文, 娄铭轩, 李沐恺, 王晗, 张亚玲. 黑龙江省水稻恶苗病致病群体[J]. 中国农业科学, 2026, 59(2): 305-321. |
| [11] | 王忠妮, 雷月, 李佳丽, 宫彦龙, 朱速松. ABC转运蛋白OsARG1调控水稻抽穗期的功能[J]. 中国农业科学, 2026, 59(1): 1-16. |
| [12] | 费耀莹, 王迪, 唐伟杰, 郭彩丽, 张小虎, 邱小雷, 程涛, 姚霞, 江冲亚, 朱艳, 曹卫星, 郑恒彪. 基于无人机多源影像融合的水稻籽粒蛋白质含量估测[J]. 中国农业科学, 2026, 59(1): 41-56. |
| [13] | 董桂春, 王子涵, 王树深, 李杰, 霍晓晴, 杨瑞, 周娟, 舒小伟, 李妍, 曹靓婧, 王子瑞, 姚友礼, 黄建晔. 硫包衣缓释肥提升水稻产量及氮肥利用率的技术途径[J]. 中国农业科学, 2026, 59(1): 57-77. |
| [14] | 王爱冬, 李瑞杰, 冯向前, 洪卫源, 李子秋, 张晓果, 王丹英, 陈松. 基于多角度成像与机器学习的水稻叶面积精确估算[J]. 中国农业科学, 2025, 58(9): 1719-1734. |
| [15] | 李云丽, 刁邓超, 刘雅睿, 孙玉晨, 孟祥宇, 邬陈芳, 汪妤, 吴建辉, 李春莲, 曾庆东, 韩德俊, 郑炜君. 小麦苗期耐热性全基因组关联分析[J]. 中国农业科学, 2025, 58(9): 1663-1683. |
|
||