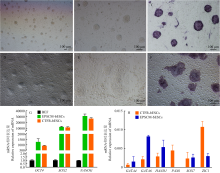
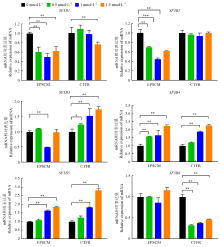
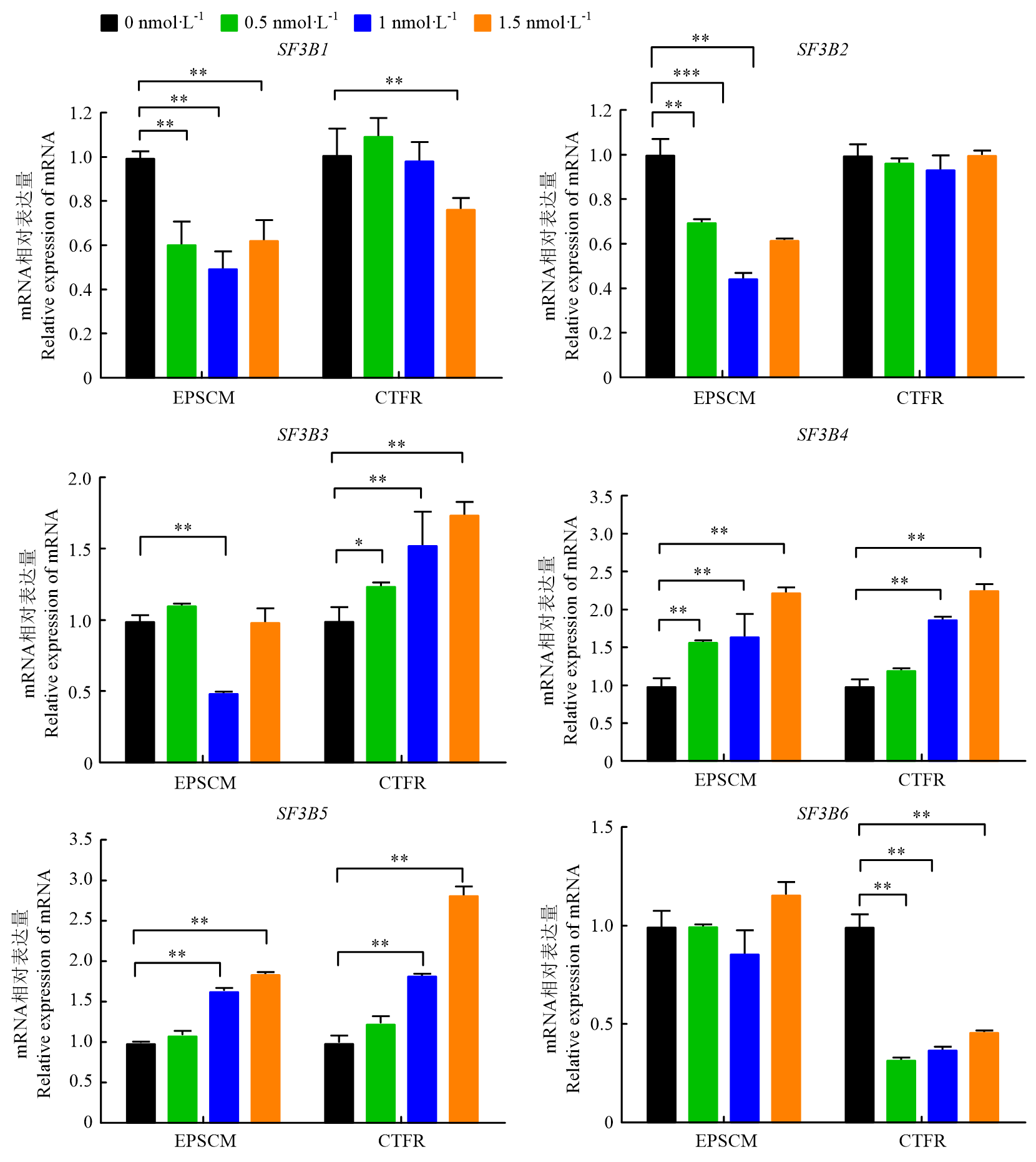
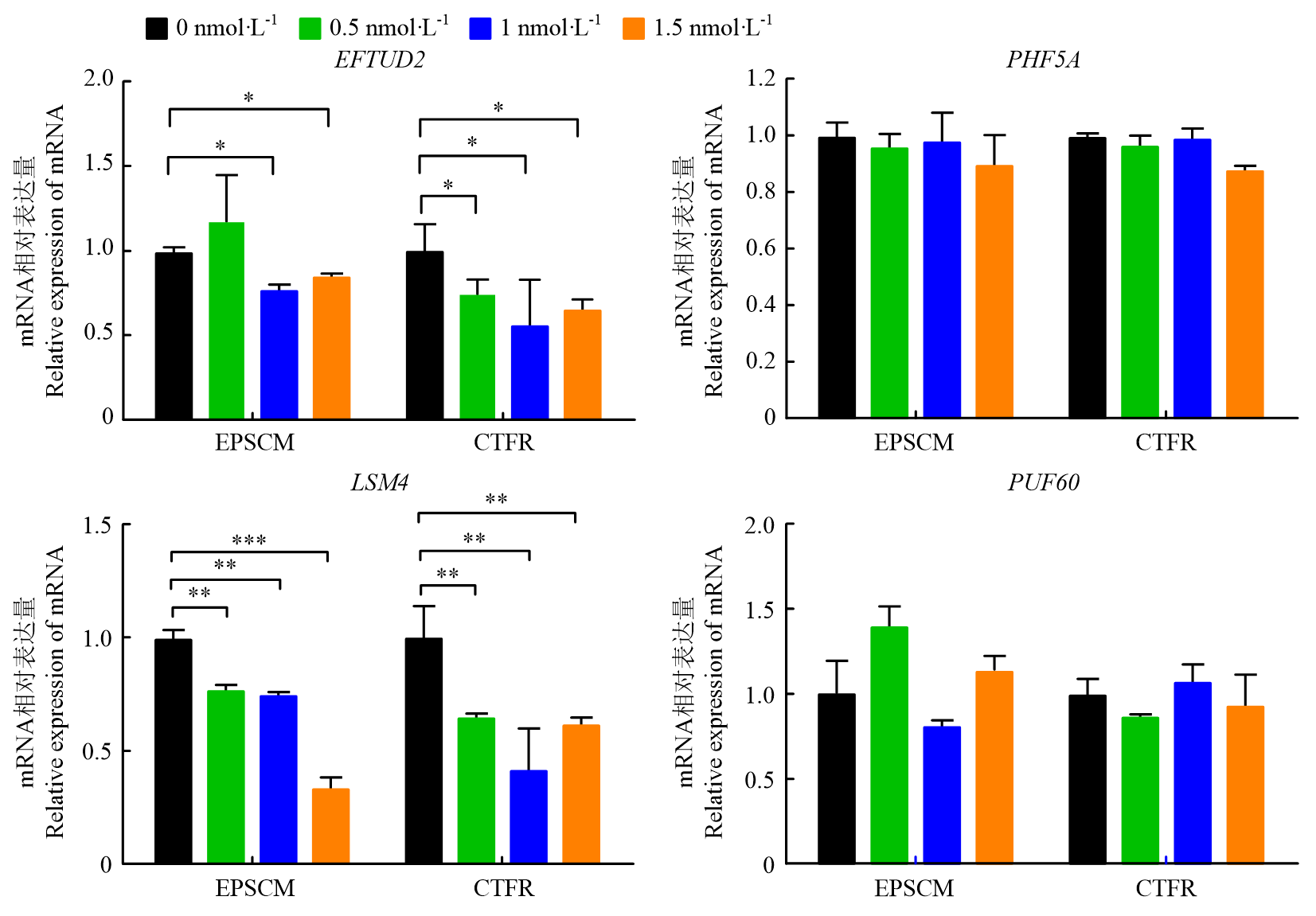
| [1] |
王冰源, 牟玉莲, 李奎, 刘志国. 农业动物干细胞研究进展. 遗传, 2020, 42(11): 1073-1080.
|
|
WANG B Y, MU Y L, LI K, LIU Z G. Research progress of stem cells in agricultural animals. Hereditas(Beijing), 2020, 42(11): 1073-1080. (in Chinese)
|
| [2] |
HOSKINS A A, RODGERS M L, FRIEDMAN L J, GELLES J, MOORE M J. Single molecule analysis reveals reversible and irreversible steps during spliceosome activation. eLife, 2016, 5: e14166.
doi: 10.7554/eLife.14166
|
| [3] |
JURICA M S, MOORE M J. Pre-mRNA splicing: awash in a sea of proteins. Molecular Cell, 2003, 12(1): 5-14.
doi: 10.1016/s1097-2765(03)00270-3
pmid: 12887888
|
| [4] |
WILL C L, LUHRMANN R. Spliceosome structure and function. Cold Spring Harbor Perspectives in Biology, 2011, 3(7): a003707.
|
| [5] |
LÓPEZ-CÁNOVAS J L, DEL RIO-MORENO M, GARCÍA- FERNANDEZ H, JIMÉNEZ-VACAS J M, MORENO-MONTILLA M T, SÁNCHEZ-FRIAS M E, AMADO V, L-LÓPEZ F, FONDEVILA M F, CIRIA R, GÓMEZ-LUQUE I, BRICEÑO J, NOGUEIRAS R, DE LA MATA M, CASTAÑO J P, RODRIGUEZ- PERÁLVAREZ M, LUQUE R M, GAHETE M D. Splicing factor SF3B1 is overexpressed and implicated in the aggressiveness and survival of hepatocellular carcinoma. Cancer Letters, 2021, 496: 72-83.
doi: 10.1016/j.canlet.2020.10.010
|
| [6] |
GÓMEZ-REDONDO I, RAMOS-IBEAS P, PERICUESTA E, FERNÁNDEZ-GONZÁLEZ R, LAGUNA-BARRAZA R, GUTIÉRREZ- ADÁN A. Minor splicing factors Zrsr1 and Zrsr2 are essential for early embryo development and 2-cell-like conversion. International Journal of Molecular Sciences, 2020, 21(11): 4115.
doi: 10.3390/ijms21114115
|
| [7] |
LÖB S, VATTAI A, KUHN C, SCHMOECKEL E, MAHNER S, WÖCKEL A, KOLBEN T, SZEKERES-BARTHO J, JESCHKE U, VILSMAIER T. Spliceosome protein EFTUD2 is upregulated in the trophoblast of spontaneous miscarriage and hydatidiform mole. Journal of reproductive immunology, 2020(140): 103149.
|
| [8] |
BOGLIOTTI Y S, WU J, VILARINO M, OKAMURA D, SOTO D A, ZHONG C Q, SAKURAI M, SAMPAIO R V, SUZUKI K, IZPISUA BELMONTE J C, ROSS P J. Efficient derivation of stable primed pluripotent embryonic stem cells from bovine blastocysts. Proceedings of the National Academy of Sciences of the United States of America, 2018, 115(9): 2090-2095.
|
| [9] |
ZHAO L X, GAO X F, ZHENG Y X, WANG Z X, ZHAO G P, REN J, ZHANG J, WU J, WU B J, CHEN Y L, SUN W, LI Y X, SU J, DING Y L, GAO Y, LIU M N, BAI X C, SUN L Z, CAO G F, TANG F C, BAO S Q, LIU P T, LI X H. Establishment of bovine expanded potential stem cells. Proceedings of the National Academy of Sciences of the United States of America, 2021(118): e2018505118.
|
| [10] |
CAPUCO A V, CHOUDHARY R K, DANIELS K M, LI R W, EVOCK-CLOVER C M. Bovine mammary stem cells: Cell biology meets production agriculture. Animal, 2012, 6(3): 382-393.
doi: 10.1017/S1751731111002369
pmid: 22436217
|
| [11] |
GUGJOO M B, AMARPAL, CHANDRA V, WANI M Y, DHAMA K, SHARMA G T. Mesenchymal stem cell research in veterinary medicine. Current Stem Cell Research & Therapy, 2018, 13(8): 645-657.
|
| [12] |
ZHAO L X, WANG Z X, ZHANG J D, YANG J, GAO X F, WU B J, ZHAO G P, BAO S Q, HU S X, LIU P T, LI X H. Characterization of the single-cell derived bovine induced pluripotent stem cells. Tissue and Cell, 2017, 49(5): 521-527.
doi: S0040-8166(17)30023-X
pmid: 28720304
|
| [13] |
PARK J W, FU S Y, HUANG B R, XU R H. Alternative splicing in mesenchymal stem cell differentiation. Stem Cells, 2020, 38(10): 1229-1240.
doi: 10.1002/stem.3248
|
| [14] |
ZHOU Z J, QU J, HE L, PENG H, CHEN P, ZHOU Y. α6-Integrin alternative splicing: Distinct cytoplasmic variants in stem cell fate specification and niche interaction. Stem Cell Research & Therapy, 2018, 9(1): 122.
|
| [15] |
SHEN H, YANG M, LI S Y, ZHANG J, PENG B, WANG C H, CHANG Z, ONG J, DU P. Mouse totipotent stem cells captured and maintained through spliceosomal repression. Cell, 2021, 184(11): 2843-2859.e20.
doi: 10.1016/j.cell.2021.04.020
pmid: 33991488
|
| [16] |
GHOSH A K, ANDERSON D D. Enantioselective total synthesis of pladienolide B: A potent spliceosome inhibitor. Organic Letters, 2012, 14(18): 4730-4733.
pmid: 22954141
|
| [17] |
YAN C Y, WAN R X, SHI Y G. Molecular mechanisms of pre-mRNA splicing through structural biology of the spliceosome. Cold Spring Harbor Perspectives in Biology, 2019, 11(1): a032409.
doi: 10.1101/cshperspect.a032409
|
| [18] |
WAHL M C, WILL C L, LÜHRMANN R. The spliceosome: Design principles of a dynamic RNP machine. Cell, 2009, 136(4): 701-718.
doi: 10.1016/j.cell.2009.02.009
pmid: 19239890
|
| [19] |
SMALL E C, LEGGETT S R, WINANS A A, STALEY J P. The EF-G-like GTPase Snu114p regulates spliceosome dynamics mediated by Brr2p, a DExD/H box ATPase. Molecular Cell, 2006, 23(3): 389-399.
pmid: 16885028
|
| [20] |
BEAUCHAMP M C, DJEDID A, DAUPIN K, CLOKIE K, KUMAR S, MAJEWSKI J, JEROME-MAJEWSKA L A. Loss of function mutation of Eftud2, the gene responsible for mandibulofacial dysostosis with microcephaly (MFDM), leads to pre-implantation arrest in mouse. PLoS ONE, 2019, 14(7): e0219280.
doi: 10.1371/journal.pone.0219280
|
| [21] |
MATERA A G, WANG Z F. A day in the life of the spliceosome. Nature Reviews Molecular Cell Biology, 2014, 15(2): 108-121. doi: 10.1038/nrm3742.
doi: 10.1038/nrm3742
pmid: 24452469
|
| [22] |
ZHOU L J, HANG J, ZHOU Y L, WAN R X, LU G F, YIN P, YAN C Y, SHI Y G. Crystal structures of the Lsm complex bound to the 3' end sequence of U6 small nuclear RNA. Nature, 2014, 506(7486): 116-120.
doi: 10.1038/nature12803
|
| [23] |
OKAMOTO K, OKAZAWA H, OKUDA A, SAKAI M, MURAMATSU M, HAMADA H. A novel octamer binding transcription factor is differentially expressed in mouse embryonic cells. Cell, 1990, 60(3): 461-472.
pmid: 1967980
|
| [24] |
PAN G J, THOMSON J A. Nanog and transcriptional networks in embryonic stem cell pluripotency. Cell Research, 2007, 17(1): 42-49.
doi: 10.1038/sj.cr.7310125
pmid: 17211451
|
| [25] |
FU X D, WU X J, DJEKIDEL M N, ZHANG Y. Myc and Dnmt1 impede the pluripotent to totipotent state transition in embryonic stem cells. Nature Cell Biology, 2019, 21(7): 835-844.
doi: 10.1038/s41556-019-0343-0
pmid: 31209294
|
| [26] |
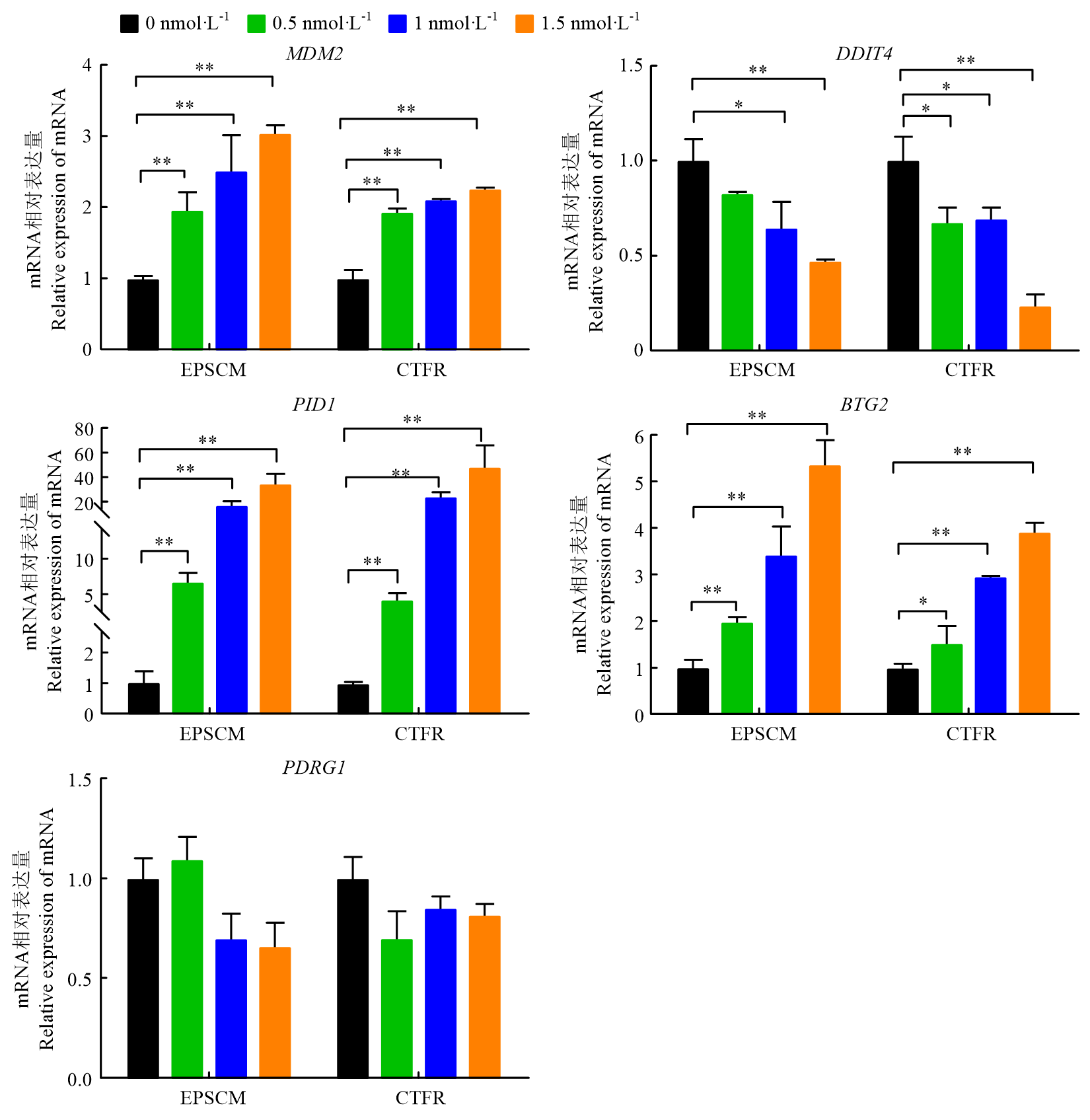
BEAUCHAMP M C, DJEDID A, BAREKE E, MERKURI F, ABER R, TAM A S, LINES M A, BOYCOTT K M, STIRLING P C, FISH J L, MAJEWSKI J, JEROME-MAJEWSKA L A. Mutation in Eftud2 causes craniofacial defects in mice via mis-splicing of Mdm2 and increased P53. Human Molecular Genetics, 2021, 30(9): 739-757.
doi: 10.1093/hmg/ddab051
|
| [27] |
WIENKEN M, MOLL U M, DOBBELSTEIN M. Mdm2 as a chromatin modifier. Journal of Molecular Cell Biology, 2016, 9(1): 74-80.
doi: 10.1093/jmcb/mjw046
|
| [28] |
HAUPT Y, MAYA R, KAZAZ A, OREN M. Mdm2 promotes the rapid degradation of p53. Nature, 1997, 387(6630): 296-299.
doi: 10.1038/387296a0
|
| [29] |
HUMPTON T J, NOMURA K, WEBER J, MAGNUSSEN H M, HOCK A K, NIXON C, DHAYADE S, STEVENSON D, HUANG D T, STRATHDEE D, BLYTH K, VOUSDEN K H. Differential requirements for MDM2 E3 activity during embryogenesis and in adult mice. Genes & Development, 2021, 35(1/2): 117-132.
doi: 10.1101/gad.341875.120
|
| [30] |
TSUI K H, CHIANG K C, LIN Y H, CHANG K S, FENG T H, JUANG H H. BTG2 is a tumor suppressor gene upregulated by p53 and PTEN in human bladder carcinoma cells. Cancer Medicine, 2018, 7(1): 184-195.
doi: 10.1002/cam4.2018.7.issue-1
|







 ), DING Qiang1, XIA ShuWen1, GAO YunDong2, LAN GuoCheng3, LIN ZhiPing4, WANG HuiLi1(
), DING Qiang1, XIA ShuWen1, GAO YunDong2, LAN GuoCheng3, LIN ZhiPing4, WANG HuiLi1(