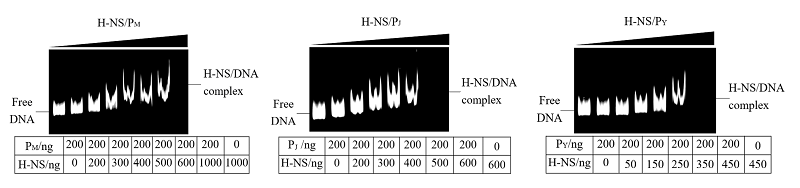
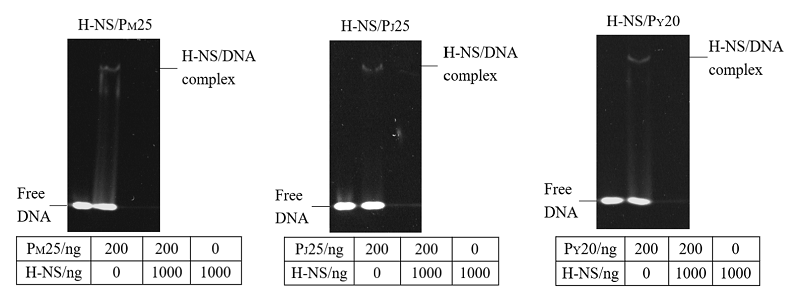
Scientia Agricultura Sinica ›› 2022, Vol. 55 ›› Issue (18): 3675-3684.doi: 10.3864/j.issn.0578-1752.2022.18.016
• ANIMAL SCIENCE·VETERINARY SCIENCE • Previous Articles
YaTing JIA( ),HuiHui HU,YaJun ZHAI,Bing ZHAO,Kun HE,YuShan PAN,GongZheng HU,Li YUAN
),HuiHui HU,YaJun ZHAI,Bing ZHAO,Kun HE,YuShan PAN,GongZheng HU,Li YUAN
| [1] |
LIU Y Y, WANG Y, WALSH T R, YI L X, ZHANG R, SPENCER J, DOI Y, TIAN G, DONG B, HUANG X, YU L F, GU D, REN H, CHEN X, LV L, HE D, ZHOU H, LIANG Z, LIU J H, SHEN J. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: a microbiological and molecular biological study. The Lancet Infectious Diseases, 2016, 16(2): 161-168. doi: 10.1016/s1473-3099(15)00424-7.
doi: 10.1016/s1473-3099(15)00424-7 |
| [2] |
WOODFORD N, CARATTOLI A, KARISIK E, UNDERWOOD A, ELLINGTON M J, LIVERMORE D M. Complete nucleotide sequences of plasmids pEK204, pEK499, and pEK516, encoding CTX-M enzymes in three major Escherichia coli lineages from the United Kingdom, all belonging to the international O25: H4-ST131 clone. Antimicrobial Agents and Chemotherapy, 2009, 53(10): 4472-4482. doi: 10.1128/aac.00688-09.
doi: 10.1128/aac.00688-09 |
| [3] |
JIAN H, XU G, GAI Y, XU J, XIAO X. The histone-like nucleoid structuring protein (H-NS) is a negative regulator of the lateral flagellar system in the deep-sea bacterium Shewanella piezotolerans WP3. Applied and Environmental Microbiology, 2016, 82(8): 2388-2398. doi: 10.1128/aem.00297-16.
doi: 10.1128/aem.00297-16 |
| [4] |
VAN DER MAAREL J R, GUTTULA D, ARLUISON V, EGELHAAF S U, GRILLO I, FORSYTH V T. Structure of the H-NS-DNA nucleoprotein complex. Soft Matter, 2016, 12(15): 3636-3642. doi: 10.1039/c5sm03076e.
doi: 10.1039/c5sm03076e |
| [5] | 徐亚珂, 龙金照, 段广才, 梁文娟, 刘慧莹, 陈帅印, 郗园林. CTX-M-15型大肠埃希菌的分子分布特征及质粒传播规律研究. 中国病原生物学杂志, 2017(11): 1048-1051. |
| XU Y K, LONG J Z, DUAN G C, LIANG W J, LIU H Y, CHEN S Y, XI Y L. The molecular distribution characteristics of and plasmid horizontal transfer in CTX-M-15-positive Escherichia coli. Journal of Pathogen Biology, 2017(11): 1048-1051. (in Chinese) | |
| [6] | PAN Y S, ZONG Z Y, YUAN L, DU X D, HUANG H, ZHONG X H, HU G Z. Complete sequence of pEC012, a multidrug-resistant IncI1 ST71 plasmid carrying blaCTX-M-65, rmtB, fosA3, floR, and oqxAB in an avian Escherichia coli ST117 strain. Frontiers in Microbiology, 2016, 7: 1117. |
| [7] |
DIACONU E L, CARFORA V, ALBA P, DI MATTEO P, STRAVINO F, BUCCELLA C, DELL'AIRA E, ONORATI R, SORBARA L, BATTISTI A, FRANCO A. Novel IncFⅡ plasmid harbouring blaNDM-4 in a carbapenem-resistant Escherichia coli of pig origin, Italy. Journal of Antimicrobial Chemotherapy, 2020, 75(12): 3475-3479.
doi: 10.1093/jac/dkaa374 |
| [8] |
XU L, WANG P, CHENG J, QIN S, XIE W. Characterization of a novel blaNDM-5-harboring IncFII plasmid and an mcr-1-bearing IncI2 plasmid in a single Escherichia coli ST167 clinical isolate. Infection and Drug Resistance, 2019, 12: 511-519. doi: 10.2147/idr.s192998.
doi: 10.2147/idr.s192998 |
| [9] |
DENG Y, ZENG Z, CHEN S, HE L, LIU Y, WU C, CHEN Z, YAO Q, HOU J, YANG T, LIU J H. Dissemination of IncFⅡ plasmids carrying rmtB and qepA in Escherichia coli from pigs, farm workers and the environment. Clinical Microbiology and Infection, 2011, 17(11): 1740-1745.
doi: 10.1111/j.1469-0691.2011.03472.x |
| [10] |
PICKER M A, WING H J. H-NS, its family members and their regulation of virulence genes in Shigella species. Genes (Basel), 2016, 7(12): 112.
doi: 10.3390/genes7120112 |
| [11] |
HÜTTENER M, PAYTUBI S, JUÁREZ A. Success in incorporating horizontally transferred genes: the H-NS protein. Trends in Microbiology, 2015, 23(2): 67-69. doi: 10.1016/j.tim.2014.12.009.
doi: 10.1016/j.tim.2014.12.009 |
| [12] |
TENDENG C, BERTIN P N. H-NS in Gram-negative bacteria: a family of multifaceted proteins. Trends in Microbiology, 2003, 11(11): 511-518. doi: 10.1016/j.tim.2003.09.005.
doi: 10.1016/j.tim.2003.09.005 |
| [13] |
RANGARAJAN A A, SCHNETZ K. Interference of transcription across H-NS binding sites and repression by H-NS. Molecular Microbiology, 2018, 108(3): 226-239. doi: 10.1111/mmi.13926.
doi: 10.1111/mmi.13926 |
| [14] |
JACQUET M, CUKIER-KAHN R, PLA J, GROS F. A thermostable protein factor acting on in vitro DNA transcription. Biochemical and Biophysical Research Communications, 1971, 45(6): 1597-1607.
doi: 10.1016/0006-291X(71)90204-X |
| [15] |
DAME R T, NOOM M C, WUITE G J. Bacterial chromatin organization by H-NS protein unravelled using dual DNA manipulation. Nature, 2006, 444(7117): 387-390.
doi: 10.1038/nature05283 |
| [16] |
DAME R T, WYMAN C, GOOSEN N. H-NS mediated compaction of DNA visualised by atomic force microscopy. Nucleic Acids Research, 2000, 28(18): 3504-10.
doi: 10.1093/nar/28.18.3504 |
| [17] |
HELGESEN E, FOSSUM-RAUNEHAUG S, SKARSTAD K. Lack of the H-NS protein results in extended and aberrantly positioned DNA during chromosome replication and segregation in Escherichia coli. Journal of Bacteriology, 2016, 198(8): 1305-1316. doi: 10.1128/jb.00919-15.
doi: 10.1128/jb.00919-15 |
| [18] |
ALI S S, SOO J, RAO C, LEUNG A S, NGAI D H, ENSMINGER A W, NAVARRE W W. Silencing by H-NS potentiated the evolution of Salmonella. PloS Pathogens, 2014, 10(11): e1004500.
doi: 10.1371/journal.ppat.1004500 |
| [19] |
SERNA A, ESPINOSA E, CAMACHO E M, CASADESÚS J. Regulation of bacterial conjugation in microaerobiosis by host- encoded functions ArcAB and sdhABCD. Genetics, 2010, 184(4): 947-958. doi: 10.1534/genetics.109.109918.
doi: 10.1534/genetics.109.109918 |
| [20] |
LU J, FROST L S. Mutations in the C-terminal region of TraM provide evidence for in vivo TraM-TraD interactions during F-plasmid conjugation. Journal of Bacteriology, 2005, 187(14): 4767-4773.
doi: 10.1128/JB.187.14.4767-4773.2005 |
| [21] |
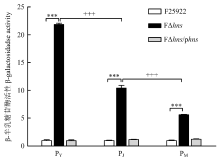
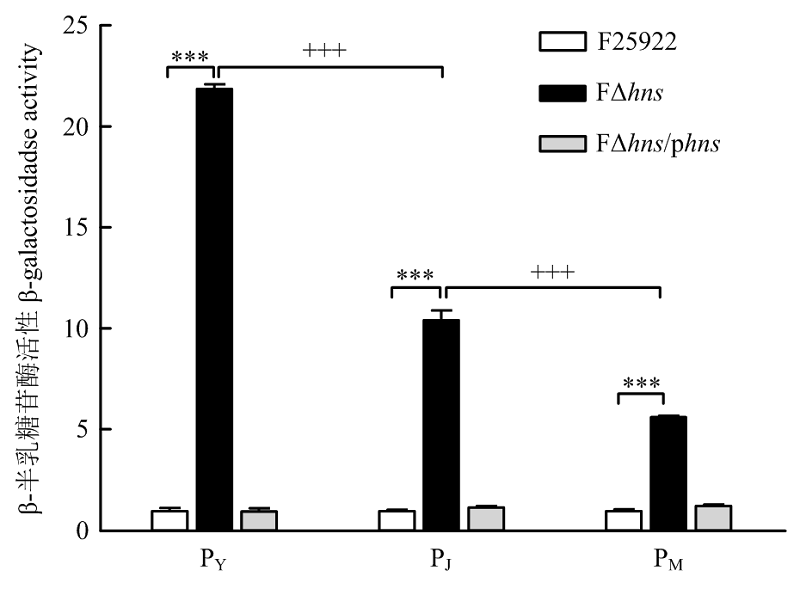
WILL W R, LU J, FROST L S. The role of H-NS in silencing F transfer gene expression during entry into stationary phase. Molecular Microbiology, 2004, 54(3): 769-782. doi: 10.1111/j.1365-2958.2004.04303.x.
doi: 10.1111/j.1365-2958.2004.04303.x. |
| [22] |
WILL W R, FROST L S. Characterization of the opposing roles of H-NS and TraJ in transcriptional regulation of the F-plasmid tra operon. Journal of Bacteriology, 2006, 188(2): 507-514. doi: 10.1128/jb.188.2.507-514.2006.
doi: 10.1128/jb.188.2.507-514.2006 |
| [23] |
PAN Y S, YUAN L, ZONG Z Y, LIU J H, WANG L F, HU G Z. A multidrug-resistance region containing blaCTX-M-65, fosA3 and rmtB on conjugative IncFⅡ plasmids in Escherichia coli ST117 isolates from chicken. Journal of Medical Microbiology, 2014, 63(Pt 3): 485-488.
doi: 10.1099/jmm.0.070664-0 |
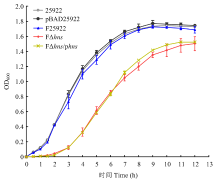
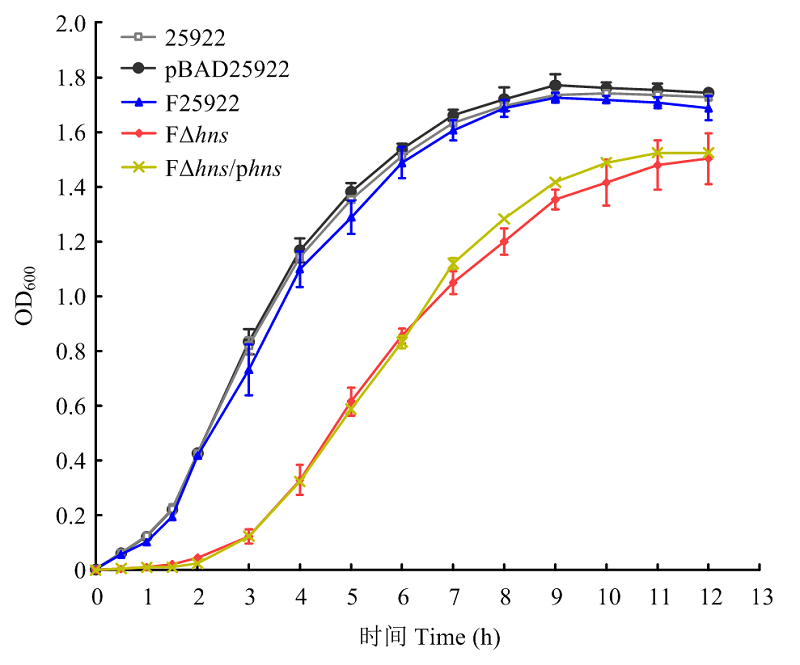
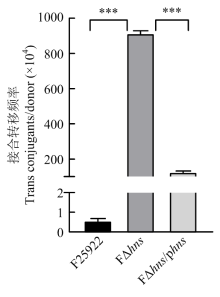
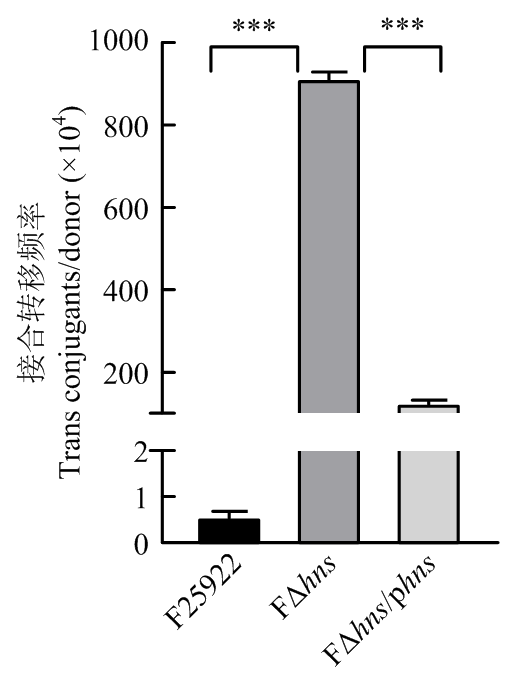
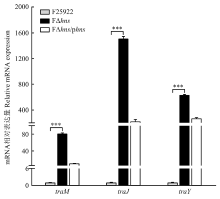
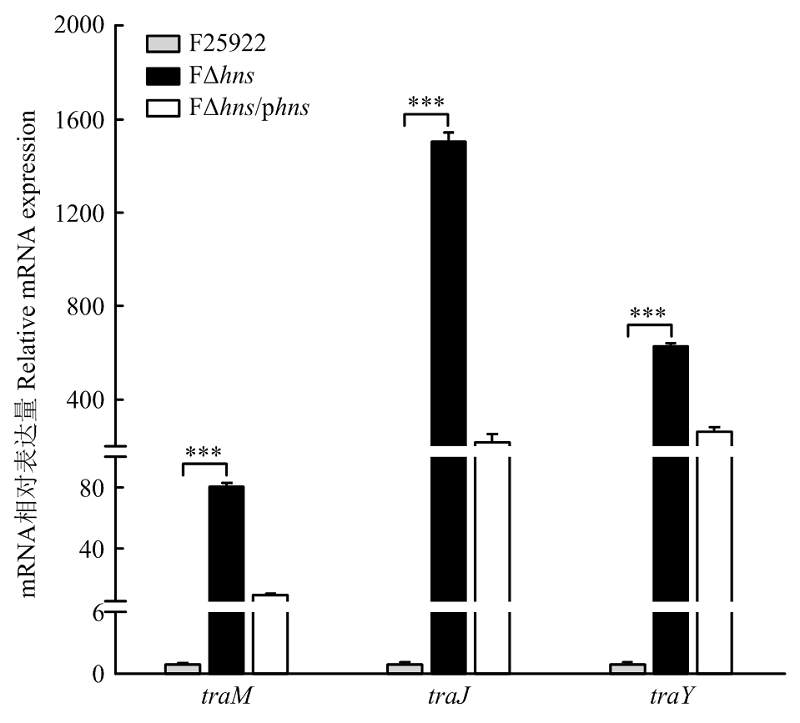
| [24] | 胡慧慧, 孙亚伟, 李文娅, 邝启红, 孙华润, 吴华, 胡功政, 苑丽. 大肠杆菌cpxR和hns双基因缺失株的构建. 中国兽医学报, 2020(1): 116-121. |
| HU H H, SUN Y W, LI W Y, KUANG Q H, SUN H R, WU H, HU G Z, YUAN L. Construction of cpxR and hns double gene deletion strain of Escherichia coli. Chinese Journal of Veterinary Science, 2020(1): 116-121. (in Chinese) | |
| [25] |
HUANG H, SUN Y, YUAN L, PAN Y, GAO Y, MA C, HU G. Regulation of the two-component regulator CpxR on aminoglycosides and β-lactams resistance in Salmonella enterica serovar Typhimurium. Frontiers in Microbiology, 2016, 7: 604. doi: 10.3389/fmicb.2016.00604.
doi: 10.3389/fmicb.2016.00604 |
| [26] |
DE LA CRUZ M A, RUIZ-TAGLE A, ARES M A, PACHECO S, YÁÑEZ J A, CEDILLO L, TORRES J, GIRÓN J A. The expression of Longus type 4 Pilus of enterotoxigenic Escherichia coli is regulated by LngR and LngS and by H-NS, CpxR and CRP global regulators. Environmental Microbiology, 2017, 19(5): 1761-1775. doi: 10.1111/1462-2920.13644.
doi: 10.1111/1462-2920.13644 |
| [27] | HUANG F, FITCHETT N, RAZO-GUTIERREZ C, LE C, MARTINEZ J, RA G, LOPEZ C, GONZALEZ L J, SIEIRA R, VILA A J, BONOMO R A, RAMIREZ M S. The H-NS regulator plays a role in the stress induced by carbapenemase expression in Acinetobacter baumannii. mSphere, 2020, 5(4): e00793-20. |
| [28] |
WILL W R, BALE D H, REID P J, LIBBY S J, FANG F C. Evolutionary expansion of a regulatory network by counter- silencing. Nature Communications, 2014, 5: 5270. doi: 10.1038/ncomms6270.
doi: 10.1038/ncomms6270 |
| [29] | LU J, PENG Y, WAN S, FROST L S, RAIVIO T, GLOVER JNM. Cooperative function of TraJ and ArcA in regulating the F plasmid tra operon. Journal of Bacteriology, 2018, 201(1): e00448-18. |
| [30] |
WAN B, ZHANG Q, TAO J, ZHOU A, YAO Y F, NI J. Global transcriptional regulation by H-NS and its biological influence on the virulence of Enterohemorrhagic Escherichia coli. Gene, 2016, 588(2): 115-123. doi: 10.1016/j.gene.2016.05.007.
doi: 10.1016/j.gene.2016.05.007 |
| [31] |
ARES M A, FERNÁNDEZ-VÁZQUEZ J L, ROSALES-REYES R, JARILLO-QUIJADA M D, VON BARGEN K, TORRES J, GONZÁLEZ-Y-MERCHAND J A, ALCÁNTAR-CURIEL M D, DE LA CRUZ M A. H-NS nucleoid protein controls virulence features of Klebsiella pneumoniae by regulating the expression of type 3 pili and the capsule polysaccharide. Frontiers in Cellular and Infection Microbiology, 2016, 6: 13. doi: 10.3389/fcimb.2016.00013.
doi: 10.3389/fcimb.2016.00013 |
| [32] |
BISCHOF K, SCHIFFER D, TRUNK S, HÖFLER T, HOPFER A, RECHBERGER G, KORAIMANN G. Regulation of R1 plasmid transfer by H-NS, ArcA, TraJ, and DNA sequence elements. Frontiers in Microbiology, 2020, 11: 1254. doi: 10.3389/fmicb.2020.01254.
doi: 10.3389/fmicb.2020.01254 |
| [1] | Jie HU,XinYi WANG,Fei WANG. Functional Characterization of BmCaspase-8-Like (BmCasp8L) as an Immune Negative Regulatory Molecule in Silkworm (Bombyx mori) [J]. Scientia Agricultura Sinica, 2018, 51(21): 4188-4196. |
| [2] | MA Xiao-Juan, LI Ya-Ming, HU Cui-Mei, WANG Fei, XIA Qing-You. Functional Characterization of BmFAF as an Immune Negative Regulatory Molecule in Silkworm (Bombyx mori) [J]. Scientia Agricultura Sinica, 2014, 47(15): 3085-3093. |
| [3] | HE Wei, WU Tian-Wen, HUANG Lei, ZHAO Wei-Min, XIA Ying, TANG Fang, MOU Yu-Lian, LI Kui. Effect of h1-Calponin Gene on in Vitro Osteogenesis Differentiation of Bone Marrow Mesenchymal Stem Cells of Wuzhishan Minipig [J]. Scientia Agricultura Sinica, 2013, 46(17): 3688-3694. |
|
||