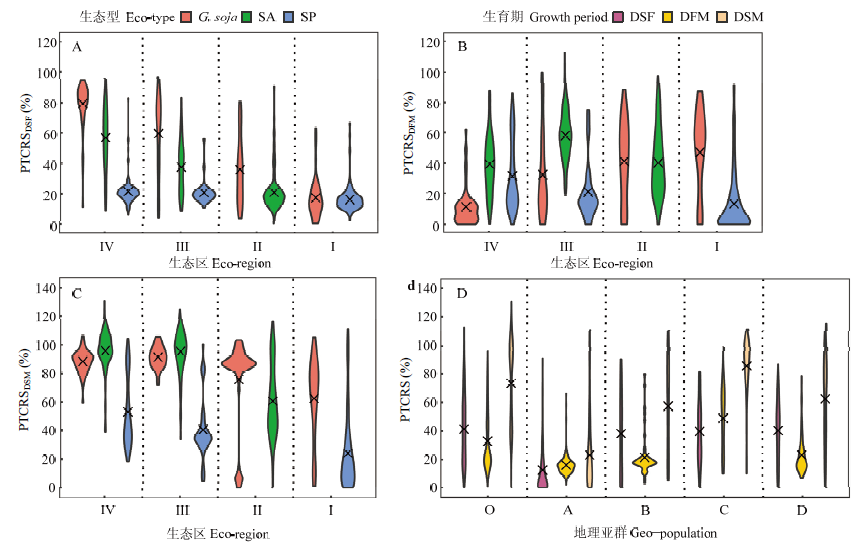
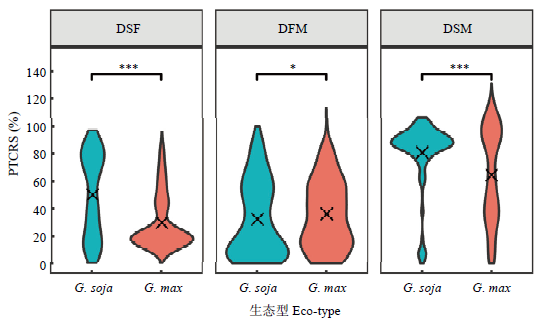
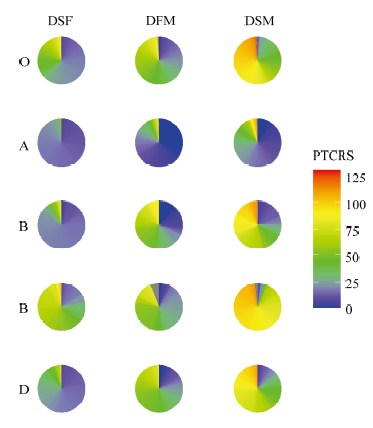
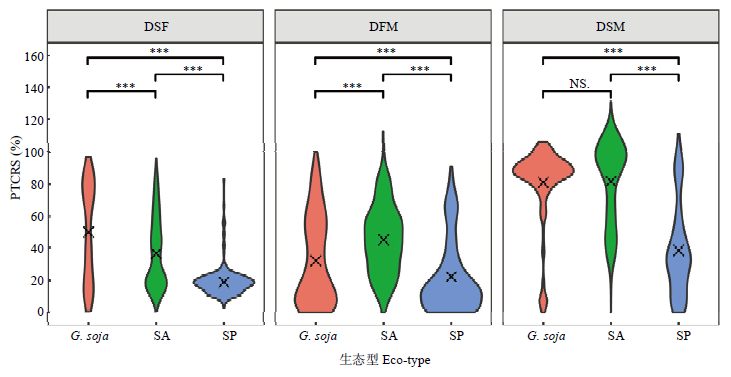
Scientia Agricultura Sinica ›› 2022, Vol. 55 ›› Issue (3): 451-466.doi: 10.3864/j.issn.0578-1752.2022.03.003
• CROP GENETICS & BREEDING·GERMPLASM RESOURCES·MOLECULAR GENETICS • Previous Articles Next Articles
JIANG FenFen( ),SUN Lei,LIU FangDong,WANG WuBin,XING GuangNan,ZHANG JiaoPing,ZHANG FengKai,LI Ning,LI Yan,HE JianBo(),GAI JunYi()
),SUN Lei,LIU FangDong,WANG WuBin,XING GuangNan,ZHANG JiaoPing,ZHANG FengKai,LI Ning,LI Yan,HE JianBo(),GAI JunYi()
| [1] |
ZHANG L X, LIU W, TSEGAW M, XU X, QI Y P, SAPEY E, LIU L P, WU T T, SUN S, HAN T F.Principles and practices of the photo-thermal adaptability improvement in soybean. Journal of Integrative Agriculture, 2020, 19(2): 295-310.
doi: 10.1016/S2095-3119(19)62850-9 |
| [2] |
COULTER M W, HAMNER K C.Photoperiodic flowering response of Biloxi soybean in 72-hour cycles. Plant Physiology, 1964, 39(5): 848-856.
doi: 10.1104/pp.39.5.848 |
| [3] | 杨永华, 盖钧镒, 马育华. 大豆生育期光温反应特性的遗传. 作物学报, 1994, 20(2): 144-148. |
| YANG Y H, GAI J Y, MA Y H.Inheritance of photothermal response characteristics in soybean growth period. Acta Agronomica Sinica, 1994, 20(2): 144-148. (in Chinese) | |
| [4] |
SONG Y H, SHIM J S, KINMONTH-SCHULTZ H A, LMAIZUMI T. Photoperiodic flowering: Time measurement mechanisms in leaves. Annual Review of Plant Biology, 2015, 66(1): 441-464.
doi: 10.1146/arplant.2015.66.issue-1 |
| [5] |
MAGALÍ N, MANTESE A I, MIRALLES D J, KANTOLIC A G.Soybean fruit development and set at the node level under combined photoperiod and radiation conditions. Journal of Experimental Botany, 2016, 67(1): 1.
doi: 10.1093/jxb/erv451 |
| [6] | 李福山. 野生大豆光温反应规律的研究. 植物遗传资源学报, 2011, 12(5): 801-805. |
| LI F S.Response regularity of wild soybean to photoperiod and temperature in natural environment. Journal of Plant Genetic Resources, 2011, 12(5): 801-805. (in Chinese) | |
| [7] | 盖钧镒, 许东河, 高忠, 岛本义也, 阿部纯, 福士泰史, 北岛俊二. 中国栽培大豆和野生大豆不同生态类型群体间遗传演化关系的研究. 作物学报, 2000, 26(5): 513-520. |
| GAI J Y, XU D H, GAO Z, SHIMAMOTO Y, ABE J, FUSHI T S, KITAJIMA S.Studies on the evolutionary relationship among ecotypes of G. max and G. soja in China. Acta Agronomica Sinica, 2000, 26(5): 513-520. (in Chinese) | |
| [8] | 范虎, 赵团结, 丁艳来, 邢光南, 盖钧镒. 中国野生大豆群体特征和地理分化的遗传分析. 中国农业科学, 2012, 45(3): 414-425. |
| FAN H, ZHAO T J, DING Y L, XING G N, GAI J Y.Genetic analysis of the characteristics and geographic differentiation of Chinese wild soybean population. Scientia Agricultura Sinica, 2012, 45(3): 414-425. (in Chinese) | |
| [41] | LÜ S L.Discussion on the origin of cultivated soybean in China. Scientia Agricultura Sinica, 1978, 11(4): 90-94. (in Chinese) |
| [9] | 朱贝贝, 孙石, 韩天富, 吴存祥. 中国不同地区野生大豆与栽培大豆生育期长度及结构性状的比较. 大豆科学, 2012, 31(6): 894-898. |
| ZHU B B, SUN S, HAN T F, WU C X.Comparison of growth period and its structure traits between wild and cultivated soybeans in China. Soybean Science, 2012, 31(6): 894-898. (in Chinese) | |
| [10] | 赵团结, 盖钧镒. 栽培大豆起源与演化研究进展. 中国农业科学, 2004, 37(7): 954-962. |
| ZHAO T J, GAI J Y.Research progress on the origin and evolution of cultivated soybean. Scientia Agricultura Sinica, 2004, 37(7): 954-962. (in Chinese) | |
| [11] |
CAI Y P, WANG L W, CHEN L, WU T T, LIU L P, SUN SH, WU C X, YAO W W, JIANG B J, YUAN S, HAN T F, HOU W S.Mutagenesis of GmFT2a and GmFT5a mediated by CRISPR/Cas9 contributes for expanding the regional adaptability of soybean. Plant Biotechnology Journal, 2020, 18(1): 298-309.
doi: 10.1111/pbi.v18.1 |
| [12] |
WU T T, LI J Y, WU C X, SUN S, MAO T T, JIANG B J, HOU W S, HAN T F.Analysis of the independent-and interactive-photo-thermal effects on soybean flowering. Journal of Integrative Agriculture, 2015, 14(4): 622-632.
doi: 10.1016/S2095-3119(14)60856-X |
| [13] |
MAO T T, LI J Y, WEN Z X, WU T T, WU C X, SUN S, JIANG B J, HOU W S, LI W B, SONG Q J, WANG D C, HAN T F.Association mapping of loci controlling genetic and environmental interaction of soybean flowering time under various photo-thermal conditions. BMC Genomics, 2017, 18(1): 415.
doi: 10.1186/s12864-017-3778-3 |
| [14] | 潘铁夫, 张德荣, 张文广, 李长荣. 东北地区大豆气候生态的研究. 吉林农业科学, 1982, 28(2): 17-28. |
| PAN T F, ZHANG D R, ZHANG W G, LI C R.Agricultural climatic ecologic factor of soybean in Northeast China. Journal of Jilin Agricultural Sciences, 1982, 28(2): 17-28. (in Chinese) | |
| [15] | 潘铁夫, 张德荣, 张文广. 东北地区大豆气候区划的研究. 大豆科学, 1983, 2(1): 1-13. |
| PAN T F, ZHANG D R, ZHANG W G.The climatic regionalization of soybean in Northeast China. Soybean Science, 1983, 2(1): 1-13. (in Chinese) | |
| [16] | 汪越胜, 盖钧镒. 中国大豆品种光温综合反应与短光照反应的关系. 中国油料作物学报, 2001, 23(2): 40-44. |
| WANG Y S, GAI J Y.The relationship between the response to short light and comprehensive responses to photo-temperature condition of soybeans from China. Chinese Journal of Oil Crop Sciences, 2001, 23(2): 40-44. (in Chinese) | |
| [17] | 费志宏, 贾贞, 冷建田, 张宝石, 吴存祥. 不同生态类型大豆品种光周期反应的鉴定. 作物杂志, 2009, 24(4): 46-49. |
| FEI Z H, JIA Z, LENG J T, ZHANG B S, WU C X.Identification of photoperiodic responses of different soybean ecotypes. Crops, 2009, 24(4): 46-49. (in Chinese) | |
| [18] |
费志宏, 吴存祥, 孙洪波, 侯文胜, 张宝石, 韩天富. 以光周期处理与分期播种试验综合鉴定大豆品种的光温反应. 作物学报, 2009, 35(8): 1525-1531.
doi: 10.3724/SP.J.1006.2009.01525 |
|
FEI Z H, WU C X, SUN H B, HOU W S, ZHANG B S, HAN T F.Identification of photothermal responses in soybean by integrating photoperiod treatments with planting-date experiments. Acta Agronomica Sinica, 2009, 35(8): 1525-1531. (in Chinese)
doi: 10.3724/SP.J.1006.2009.01525 |
|
| [19] | GARNER W W, ALLARD H A.Further studies in photoperiodism: the response of the plant to relative length of day and night. Journal of Agricultural Research, 1923, 23: 871-920. |
| [20] |
GARNER W W, ALLARD H A.Comparative responses of long-day and short-day plants to relative length of day and night. Plant Physiology, 1933, 8(3): 347-356.
doi: 10.1104/pp.8.3.347 |
| [21] |
BORTHWICK H A, PARKER M W.Photoperiodic responses of several varieties of soybeans. Botanical Gazette, 1939, 101(2): 341-365.
doi: 10.1086/334874 |
| [22] | UPADHYAY A P, SUMMERFIELD R H, ELLIS R H, ROBERTS R H, QI A.Variation in the durations of the photoperiod-sensitive and photoperiod-insensitive phases of development to flowering among eight maturity isolines of soyabean [Glycine max (L.) Merrill]. Annual Botany, 1994, 74(1): 97-101. |
| [23] | 韩天富. 大豆开花后的光周期反应. 大豆科学, 1996, 15(1): 69-73. |
| HAN T F.A review on the post-flowering photoperiodic responses in soybean. Soybean Science, 1996, 15(1): 69-73. (in Chinese) | |
| [24] | 韩天富, 盖钧镒, 邱家驯. 中国大豆不同生态类型代表品种开花前、开花后光周期反应的比较研究. 大豆科学, 1998, 17(2): 35-40. |
| HAN T F, GAI J Y, QIU J X.A comparative study on pre-and post-flowering photoperiod response in various ecotypes of soybeans. Soybean Science, 1998, 17(2): 35-40. (in Chinese) | |
| [25] | RAHMAN M M, HAMPTON J G, HILL M J.Soybean development under the cool temperate environment of Canterbury, New Zealand. Journal of New Seeds, 2006, 7(4): 17-36. |
| [26] |
KANTOLIC A G, SLAFER G A.Development and seed number in indeterminate soybean as affected by timing and duration of exposure to long photoperiods after flowering. Annuals of Botany, 2007, 99(5): 925-933.
doi: 10.1093/aob/mcm033 |
| [27] | 徐豹, 路琴华. 大豆生态研究: Ⅰ.中国不同纬度野生大豆的光温生态分析. 大豆科学, 1983, 2(3): 155-168. |
| XU B, LU Q H.Research on soybean ecology: I. ecological analysis of light and temperature of wild soybeans in different latitudes in China. Soybean Science, 1983, 2(3): 155-168. (in Chinese) | |
| [28] | 汪越胜, 马宏惠. 中国大豆地理生态型生育前期光温综合反应. 安徽师范大学学报(自然科学版), 2000, 28(1): 40-42. |
| WANG Y S, MA H H.Photothermal comprehensive response of days to flowering of soybean ecotypes of China. Journal of Anhui Normal University (Natural Science Edition), 2000, 28(1): 40-42. (in Chinese) | |
| [29] | 汪越胜, 陈冬生, 马宏惠. 中国大豆成熟期光温综合反应与短光照反应间关系. 安徽师范大学学报(自然科学版), 2000, 23(3): 231-233. |
| WANG Y S, CHEN D S, MA H H.Relationship between photothermal comprehensive response and response to short photoperiod of Chinese soybeans in days to maturity. Journal of Anhui Normal University (Natural Science Edition), 2000, 23(3): 231-233. (in Chinese) | |
| [30] |
SINGH R J, HYMOWITZ T.Soybean genetic resources and crop improvement. Genome, 1999, 42(4): 605-616.
doi: 10.1139/g99-039 |
| [31] |
LIU X Q, WU J A, REN H X, QI Y X, LI C Y, CAO J Q, ZHANG X Y, ZHANG Z P, CAI Z Y, GAI J Y.Genetic variation of world soybean maturity date and geographic distribution of maturity groups. Breeding Science, 2017, 67(3): 221-232.
doi: 10.1270/jsbbs.16167 |
| [32] | 徐豹. 中国野生大豆(G. soja)研究十年.吉林农业科学, 1989, 55(1): 5-13. |
| XU B.Ten years of research on Chinese wild soybeans (G. soja). Journal of Jilin Agricultural Sciences, 1989, 55(1): 5-13. (in Chinese) | |
| [33] | 李莹. 野生大豆和生态环境关系的统计分析. 中国油料, 1981, 3(4): 57-60. |
| LI Y.Statistical analysis of the relationship between wild soybean and ecological environment. Chinese Journal of Oil Crop Sciences, 1981, 3(4): 57-60. (in Chinese) | |
| [34] | 徐树传, 童川拉, 游榕青, 陈孝宽, 危嫩弟. 野生大豆光温反应观察研究. 福建农业科技, 1985, 16(5): 19-20. |
| XU S Z, TONG C L, YOU R Q, CHEN X K, WEI N D.Observation and research on the photothermal response of wild soybean. Fujian Agricultural Science and Technology, 1985, 16(5): 19-20. (in Chinese) | |
| [35] | 舒世珍, 李福山, 常汝镇. 大豆主要性状演化的初步研究. 作物学报, 1986, 12(4): 255-260. |
| SHU S Z, LI F S, CHANG R Z.Preliminary study on the evolution of soybean main traits. Acta Agronomica Sinica, 1986, 12(4): 255-260. (in Chinese) | |
| [36] | 徐豹, 路琴华. 不同进化类型大豆花荚形成和脱落的比较研究. 大豆科学, 1988, 7(2): 103-112. |
| XU B, LU Q H.Comparative study on the formation and shedding of soybean flower pods with different evolution types. Soybean Science, 1988, 7(2): 103-112. (in Chinese) | |
| [37] | 韩天富, 盖钧镒, 陈风云, 邱家驯. 生育期结构不同的大豆品种的光周期反应和农艺性状. 作物学报, 1998, 24(5): 550-557. |
| HAN T F, GAI J Y, CHEN F Y, QIU J X.Photoperiod response and agronomic traits of soybean varieties with different growth period structures. Acta Agronomica Sinica, 1998, 24(5): 550-557. (in Chinese) | |
| [38] | 常汝镇. 关于栽培大豆起源的研究. 中国油料, 1989, 11(1): 3-9. |
| CHANG R Z.Research on the origin of cultivated soybean. Chinese Journal of Oil Crop Sciences, 1989, 11(1): 3-9. (in Chinese) | |
| [39] |
WANG L X, GUAN R X, LIU Z X, CHANG R Z, QIU L J.Genetic diversity of Chinese cultivated soybean revealed by SSR markers. Crop Science, 2006, 46(3): 1032-1038.
doi: 10.2135/cropsci2005.0051 |
| [40] |
LI Y H, ZHANG C,SMULDERS M J M, LI W, MA Y S, XU Q, CHANG R Z, QIU L J. Analysis of average standardized SSR allele size supports domestication of soybean along the Yellow River. Genetic Resources and Crop Evolution, 2013, 60(2): 763-776.
doi: 10.1007/s10722-012-9873-z |
| [41] | 吕世霖. 关于我国栽培大豆原产地问题的探讨. 中国农业科学, 1978, 11(4): 90-94. |
| No related articles found! |
|
||