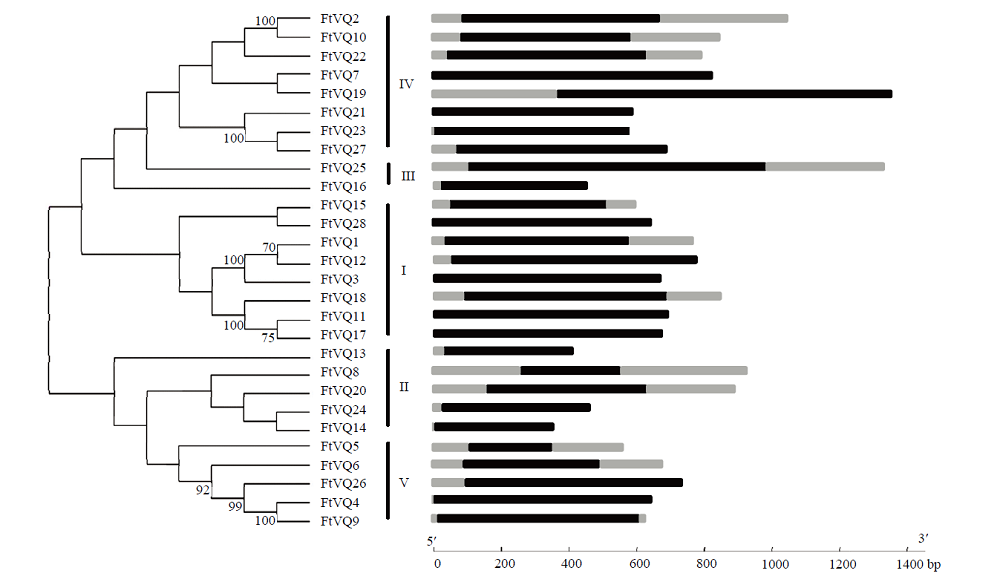
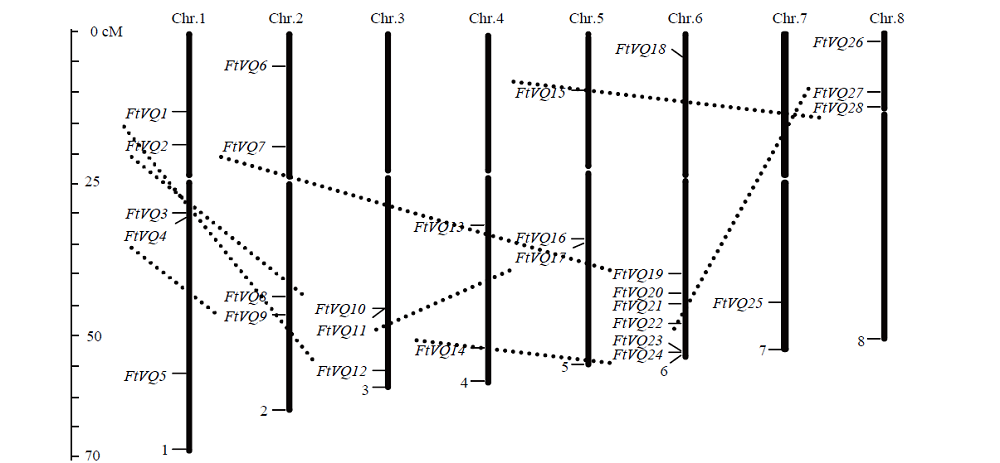
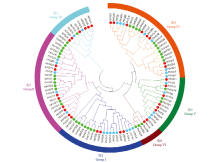
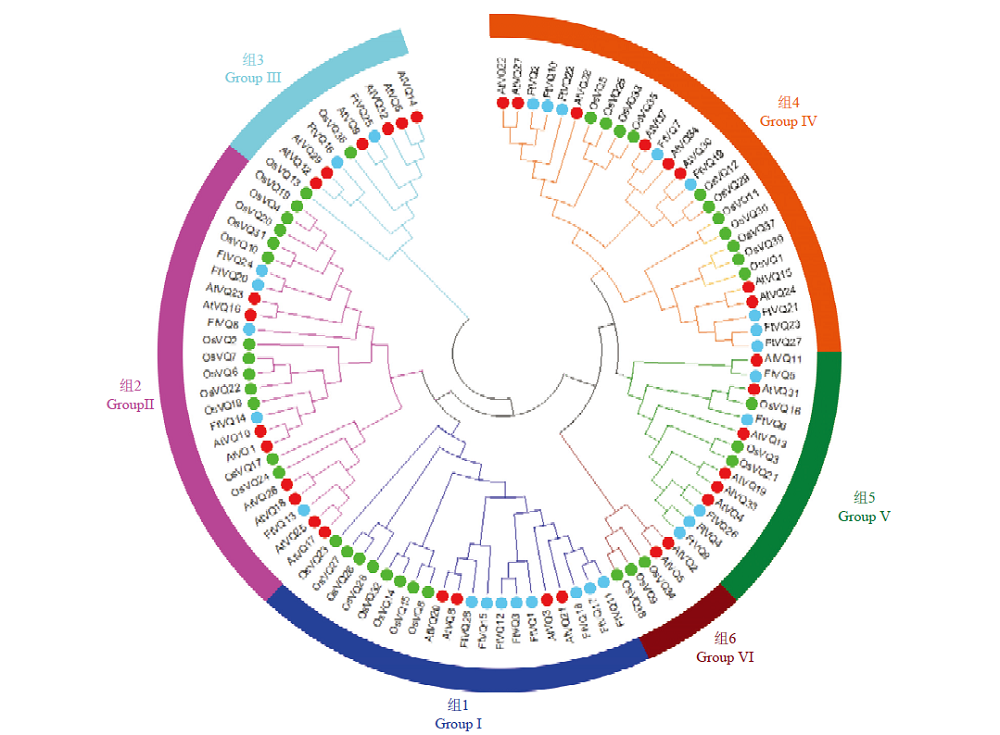
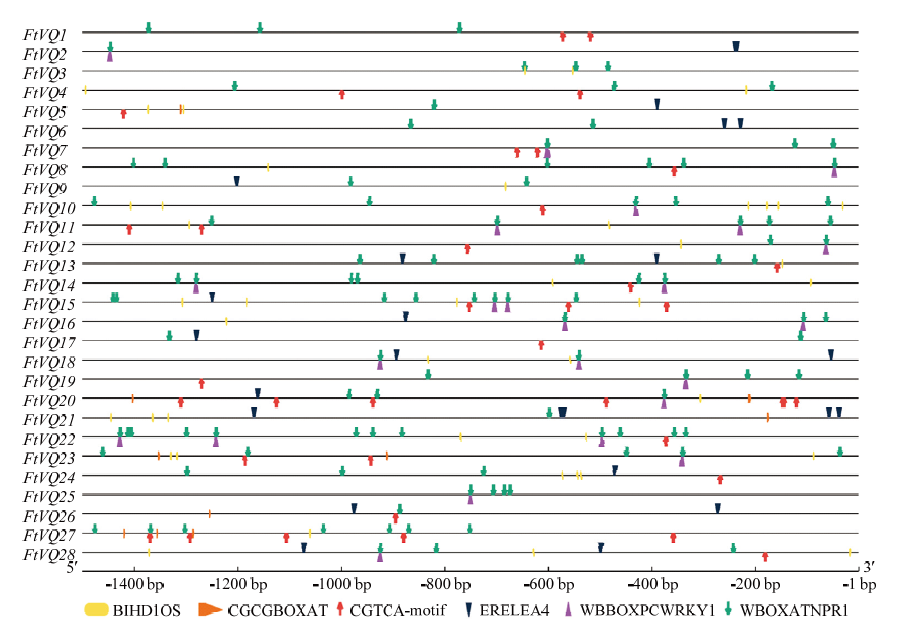
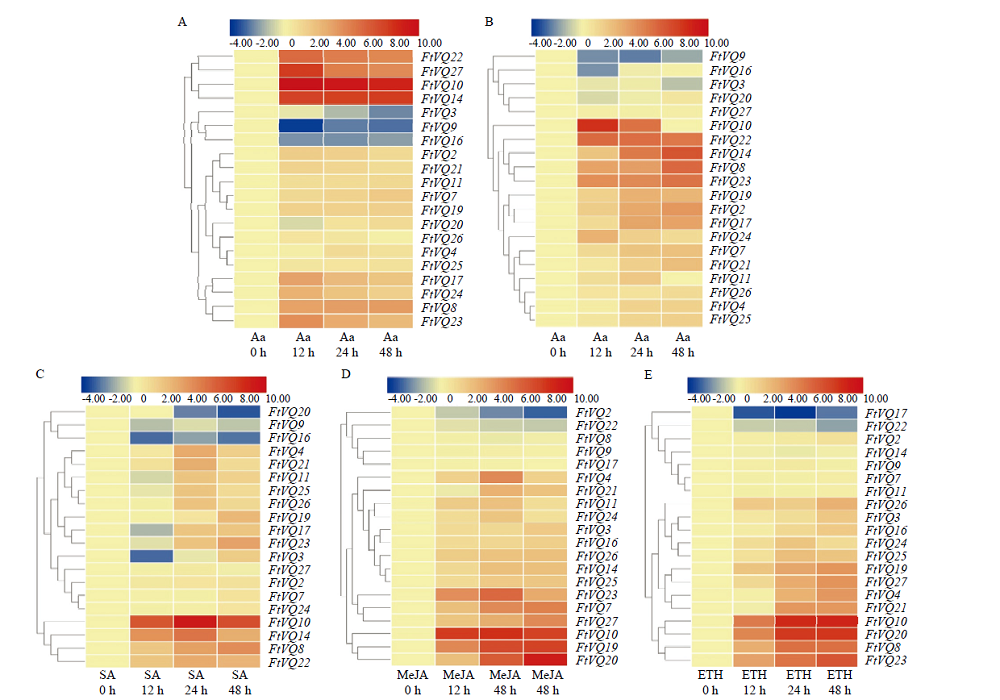
Scientia Agricultura Sinica ›› 2021, Vol. 54 ›› Issue (19): 4048-4060.doi: 10.3864/j.issn.0578-1752.2021.19.002
• CROP GENETICS & BREEDING·GERMPLASM RESOURCES·MOLECULAR GENETICS • Previous Articles Next Articles
ZHENG FengSheng1( ),WANG HaiHua1,2(),WU QingTao1,SHEN Quan1,2,TIAN JianHong1,PENG XiXu1,3,TANG XinKe1,3()
),WANG HaiHua1,2(),WU QingTao1,SHEN Quan1,2,TIAN JianHong1,PENG XiXu1,3,TANG XinKe1,3()
| [1] |
CHI Y, YANG Y, ZHOU Y, ZHOU J, FAN B, YU J Q, CHEN Z. Protein-protein interactions in the regulation of WRKY transcription factors. Molecular Plant, 2013, 6(2):287-300.
doi: 10.1093/mp/sst026 |
| [2] |
JING Y, LIN R. The VQ motif-containing protein family of plant- specific transcriptional regulators. Plant Physiology, 2015, 169(1):371-378.
doi: 10.1104/pp.15.00788 |
| [3] |
CHENG Y, ZHOU Y, YANG Y, CHI Y J, ZHOU J, CHEN J Y, WANG F, FAN B, SHI K, ZHOU Y H, YU J Q, CHEN Z. Structural and functional analysis of VQ motif-containing proteins in Arabidopsis as interacting proteins of WRKY transcription factors. Plant Physiology, 2012, 159(2):810-825.
doi: 10.1104/pp.112.196816 |
| [4] | WEYHE M, ESCHEN-LIPPOLD L, PECHER P, SCHEEL D, LEE J. Ménage à trois: The complex relationships between mitogen- activated protein kinases, WRKY transcription factors, and VQ-motif- containing proteins. Plant Signaling & Behavior, 2014, 9(8):29519. |
| [5] |
ZHANG L, LI X, MA B, GAO Q, DU H, HAN Y, LI Y, CAO Y, QI M, ZHU Y, LU H, MA M, LIU L, ZHOU J, NAN C, QIN Y, WANG J, CUI L, LIU H, LIANG C, QIAO Z. The tartary buckwheat genome provides insights into rutin biosynthesis and abiotic stress tolerance. Molecular Plant, 2017, 10(9):1224-1237.
doi: 10.1016/j.molp.2017.08.013 |
| [6] |
YANG J, DUAN G, LI C, LIU L, HAN G, ZHANG Y, WANG C. The crosstalks between jasmonic acid and other plant hormone signaling highlight the involvement of jasmonic acid as a core component in plant response to biotic and abiotic stresses. Frontiers in Plant Science, 2019, 10:1349.
doi: 10.3389/fpls.2019.01349 |
| [7] |
KIM DY, KWON S I, CHOI C, LEE H, AHN I, PARK S R, BAE S C, LEE S C, HWANG D J. Expression analysis of rice VQ genes in response to biotic and abiotic stresses. Gene, 2013, 529(2):208-214.
doi: 10.1016/j.gene.2013.08.023 |
| [8] |
LI N, LI X, XIAO J, WANG S. Comprehensive analysis of VQ motif-containing gene expression in rice defense responses to three pathogens. Plant Cell Report, 2014 33(9):1493-1505.
doi: 10.1007/s00299-014-1633-4 |
| [9] | ZHOU Y, YANG Y, ZHOU X, CHI Y, FAN B, CHEN Z. Structural and functional characterization of the VQ protein family and VQ protein variants from soybean. Scientific Report, 2016, 6:34663. |
| [10] |
LIU C, LIU H, ZHOU C, TIMKO M P. Genome-wide identification of the VQ protein gene family of tobacco (Nicotiana tabacum L.) and analysis of its expression in response to phytohormones and abiotic and biotic stresses. Genes, 2020, 11(3):284.
doi: 10.3390/genes11030284 |
| [11] |
LAI Z, LI Y, WANG F, CHENG Y, FAN B, YU J Q, CHEN Z. Arabidopsis sigma factor binding proteins are activators of the WRKY33 transcription factor in plant defense. The Plant Cell, 2011, 23(10):3824-3841.
doi: 10.1105/tpc.111.090571 |
| [12] | WANG H, HU Y, PAN J, YU D. Arabidopsis VQ motif-containing proteins VQ12 and VQ29 negatively modulate basal defense against Botrytis cinerea. Scientific Report, 2015, 5:14185. |
| [13] |
ALI M R M, UEMURA T, RAMADAN A, ADACHI K, NEMOTO K, NOZAWA A, HOSHINO R, ABE H, SAWASAKI T, ARIMURA G I. The ring-type E3 ubiquitin ligase JUL1 targets the VQ-motif protein JAV1 to coordinate jasmonate signaling. Plant Physiology, 2019, 179(4):1273-1284.
doi: 10.1104/pp.18.00715 |
| [14] | CHU W Y, LIU B, WANG Y J, PAN F, CHEN Z, YAN H W, XIANG Y. Genome-wide analysis of poplar VQ gene family and expression profiling under PEG, NaCl, and SA treatments. Tree Genetics & Genomes, 2016, 12:124. |
| [15] | SONG W, ZHAO H, ZHANG X, LEI L, LAI J. Genome-wide identification of VQ motif-containing proteins and their expression profiles under abiotic stresses in maize. Frontiers in Plant Science, 2016, 6:1177. |
| [16] |
ZHANG G, WANG F, LI J, DING Q, ZHANG Y, LI H, ZHANG J, GAO J. Genome-wide identification and analysis of the VQ motif-containing protein family in Chinese cabbage (Brassica rapa L. ssp. Pekinensis). International Journal of Molecular Sciences, 2015, 16(12):28683-28704.
doi: 10.3390/ijms161226127 |
| [17] | WANG M, VANNOZZI A, WANG G, ZHONG Y, CORSO M, CAVALLINI E, CHENG Z M. A comprehensive survey of the grapevine VQ gene family and its transcriptional correlation with WRKY proteins. Frontiers in Plant Science, 2015, 6:417. |
| [18] |
CHEN P, WEI F, CHENG S, MA L, WANG H, ZHANG M, MAO G, LU J, HAO P, AHMAD A, GU L, MA Q, WU A, WEI H, YU S. A comprehensive analysis of cotton VQ gene superfamily reveals their potential and extensive roles in regulating cotton abiotic stress. BMC Genomics, 2020, 21(1):795.
doi: 10.1186/s12864-020-07171-z |
| [19] | GARRIDO-GALA J, HIGUERA J J, MUÑOZ-BLANCO J, AMIL-RUIZ F, CABALLERO J L. The VQ motif-containing proteins in the diploid and octoploid strawberry. Scientific Report, 2019, 9(1):4942. |
| [20] |
WANG Y, LIU H, ZHU D, GAO Y, YAN H, XIANG Y. Genome-wide analysis of VQ motif-containing proteins in Moso bamboo (Phyllostachys edulis). Planta, 2017, 246(1):165-181.
doi: 10.1007/s00425-017-2693-9 |
| [21] | LING L, QU Y, ZHU J, WANG D, GUO C. Genome-wide identification and expression analysis of the VQ gene family in Cicer arietinum and Medicago truncatula. PeerJ, 2020, 8:8471. |
| [22] |
SHEN Q, PENG X X, HE F, LI S Q, XIAO Z Y, WANG H H, TANG X K, ZHOU M L. First report of Nigrospora osmanthi causing leaf spot on tartary buckwheat in China. Plant Disease, 2020. doi: 10.1094/PDIS-08-20-1773-PDN.
doi: 10.1094/PDIS-08-20-1773-PDN |
| [23] |
LYNCH M, CONERY J S. The evolutionary fate and consequences of duplicate genes. Science, 2000, 290(5494):1151-1155.
doi: 10.1126/science.290.5494.1151 |
| [24] |
LI W H, GOJOBORI T, NEI M. Pseudogenes as a paradigm of neutral evolution. Nature, 1981, 292:237-239.
doi: 10.1038/292237a0 |
| [25] |
STORZ J F. Genome evolution: Gene duplication and the resolution of adaptive conflict. Heredity, 2009, 102(2):99-100.
doi: 10.1038/hdy.2008.114 |
| [26] |
ZHANG G, WEI B. Characterization of VQ motif-containing protein family and their expression patterns under phytohormones and abiotic stresses in melon (Cucumis melo L.). Plant Growth Regulation, 2019, 89(3):273-285.
doi: 10.1007/s10725-019-00534-x |
| [27] |
JIANG S Y, SEVUGAN M, RAMACHANDRAN S. Valine-glutamine (VQ) motif coding genes are ancient and non-plant-specific with comprehensive expression regulation by various biotic and abiotic stress. BMC Genomics, 2018, 19:342.
doi: 10.1186/s12864-018-4733-7 |
| [28] |
UJI Y, KASHIHARA K, KIYAMA H, MOCHIZUKI S, AKIMITSU K, GOMI K. Jasmonic acid-induced VQ-motif-containing protein OsVQ13 influences the OsWRKY45 signaling pathway and grain size by associating with OsMPK6 in rice. International Journal of Molecular Science, 2019, 20:2917.
doi: 10.3390/ijms20122917 |
| [29] |
EULGEM T, RUSHTON P J, ROBATZEK S, SOMSSICH I E. The WRKY superfamily of plant transcription factors. Trends in Plant Science, 2000, 5(5):199-206.
doi: 10.1016/S1360-1385(00)01600-9 |
| No related articles found! |
|
||