






中国农业科学 ›› 2026, Vol. 59 ›› Issue (2): 368-385.doi: 10.3864/j.issn.0578-1752.2026.02.011
王峰1,2( ), 常云妮1,2, 吴志丹1,2, 孙君1,2, 江福英1,2, 陈玉真1,2(), 余文权1,3()
), 常云妮1,2, 吴志丹1,2, 孙君1,2, 江福英1,2, 陈玉真1,2(), 余文权1,3()
收稿日期:2025-03-10
接受日期:2025-07-17
出版日期:2026-01-16
发布日期:2026-01-22
通信作者:
联系方式:
王峰,E-mail:82458lin@163.com。
基金资助:
WANG Feng1,2(), CHANG YunNi1,2, WU ZhiDan1,2, SUN Jun1,2, JIANG FuYing1,2, CHEN YuZhen1,2(), YU WenQuan1,3()
Received:2025-03-10
Accepted:2025-07-17
Published:2026-01-16
Online:2026-01-22
摘要:
【目的】 真菌是调控土壤生态功能的重要因素,解析长期施氮条件下茶园土壤真菌群落多样性、功能类群及群落构建机制的变化规律,确定茶园氮肥适宜用量,为制定茶园长期合理施氮量提供科学依据。 【方法】 依托于2011年开始的长期田间水泥池定位试验,设置4种施氮水平(不施氮肥,N0;低量施氮,112.5 kg·hm-2,N1;中量施氮,225 kg·hm-2,N2;高量施氮,450 kg·hm-2,N3),采集第10年(2020年)春茶和秋茶季节土壤样品,利用ITS高通量测序技术分析长期施氮对土壤真菌多样性、功能类群及群落构建的影响。 【结果】 与N0处理相比,长期施氮均可显著提高茶鲜叶产量,增幅分别是137.8%—430.2%(春季)和33.4%—67.5%(秋季),其中N2和N3处理差异不显著。随着施氮量的增加,真菌多样性呈先增加后降低趋势,N3处理真菌Ace、Chao1和Shannon指数显著低于N0处理。非度量多维度分析(NMDS)和置换多元方差分析(PERMANOVA)结果表明,施氮显著改变真菌的群落结构。FUNGuild真菌功能预测发现,长期施氮显著改变春茶期间土壤真菌功能,秋茶则变化不显著。与N0处理相比,N2和N3处理降低茶园土壤植物病原菌和土壤腐生真菌的相对丰度(尤其是春茶),N2处理提高了土壤欧石南类菌根和丛枝菌根两类有益真菌的相对丰度。冗余分析结果表明,土壤pH、硝态氮、铵态氮和速效钾是影响土壤真菌群落结构和功能类群变化最主要的因素。与N0处理相比,施氮处理均增加了真菌网络边数、平均度、平均聚类系数和网络密度,增强了网络结构稳定性。在群落演替过程中,随机性过程主导长期施氮下茶园土壤真菌群落构建,中低氮处理增强了确定性过程在群落组装中的作用。 【结论】 从茶叶产量、群落多样性、富集有益微生物及网络稳定性角度综合分析,施用氮肥225 kg·hm-2是较合理的氮肥用量。
王峰, 常云妮, 吴志丹, 孙君, 江福英, 陈玉真, 余文权. 长期施氮对茶园土壤真菌多样性、功能类群及群落构建的影响[J]. 中国农业科学, 2026, 59(2): 368-385.
WANG Feng, CHANG YunNi, WU ZhiDan, SUN Jun, JIANG FuYing, CHEN YuZhen, YU WenQuan. Effects of Long-Term Nitrogen Application on Soil Fungal Diversity, Functional Groups and Assembly Processes in Tea Gardens[J]. Scientia Agricultura Sinica, 2026, 59(2): 368-385.
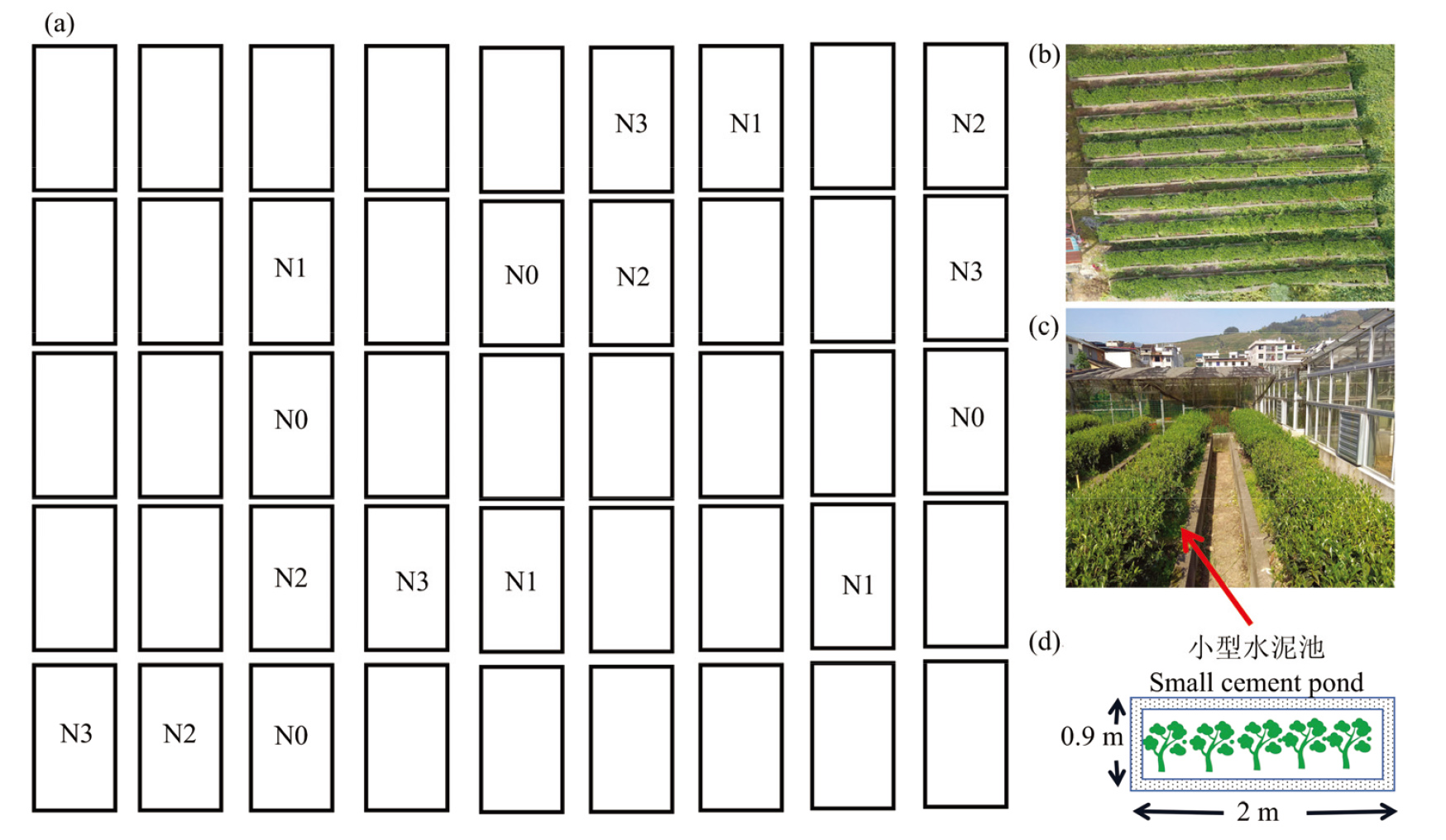
图1
茶园长期施氮试验设置 a:田间每个独立的小水泥池(空白处为其他试验处理);b:田间航拍效果图;c:用水泥池隔离的小区;d:每个小区的尺寸图"
表1
不同施氮处理下茶园土壤理化性质和茶叶产量"
| 指标 Indicator | 春季 Spring | 秋季 Autumn | 施氮量 Nitrogen (N) | 季节 Seasons(S) | N×S | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| N0 | N1 | N2 | N3 | N0 | N1 | N2 | N3 | ||||
| SOM (g·kg-1) | 19.02±2.67Ab | 20.14±1.58Aab | 19.67±1.19Aab | 22.38±1.46Aa | 15.53±3.11Aa | 21.07±1.98Aa | 17.34±3.23Aa | 31.5±11.69Ab | 6.792*** | 0.412NS | 2.98NS |
| TN (g·kg-1) | 0.69±0.04Bb | 0.89±0.09Aa | 0.96±0.05Aa | 0.97±0.13Aa | 0.87±0.06Ab | 1.04±0.11Aa | 0.97±0.17Aa | 1.44±0.22Aa | 14.465*** | 19.718*** | 4.231* |
| AN (mg·kg-1) | 29.03±5.44Ab | 35.93±8.29Aa | 40.22±9.91Aa | 58.18±7.45Aa | 39.90±3.54Ab | 40.93±11.57Ab | 42.97±0.05Ab | 62.93±10.48Aa | 16.835*** | 4.296 | 0.386NS |
| NO3--N (mg·kg-1) | 3.48±0.58Ac | 2.13±2.09Ac | 9.48±2.71Ab | 14.53±5.04Aa | 1.04±0.40Ab | 1.27±0.23Ab | 1.29±0.17Ab | 2.38±0.81Bb | 16.223*** | 58.105*** | 11.327*** |
| NH4+-N (mg·kg-1) | 13.4±5.38Bb | 19.19±1.46Bb | 25.01±7.86Bb | 40.98±17.52Aa | 50.54±0.97Aa | 51.19±0.69Aa | 50.12±0.77Aa | 50.32±1.23Aa | 5.403** | 106.334*** | 5.789** |
| TP (g·kg-1) | 0.60±0.05Aa | 0.71±0.11Aa | 0.63±0.12Aa | 0.68±0.02Aa | 0.61±0.03Ab | 0.65±0.09Ab | 0.67±0.03Ab | 0.76±0.04Aa | 3.189* | 0.524NS | 1.077NS |
| AP (mg·kg-1) | 73.87±18.79Ba | 61.34±16.14Aa | 76.3±37.53Aa | 80.75±3.57Ba | 183.76±37.44Aa | 91.24±41.03Ab | 85.22±50.91Ab | 197.17±62.2Aa | 5.758** | 24.271*** | 4.157* |
| TK (g·kg-1) | 28.02±1.51Aa | 27.66±0.75Aa | 25.97±2.04Aa | 29.17±1.44Aa | 28.07±1.34Aab | 26.9±0.92Ab | 26.47±0.97Ab | 29.42±0.78Aa | 8.027*** | 0.01NS | 0.36NS |
| AK (mg·kg-1) | 291.01±21.9Aa | 303.66±7.13Aa | 243.01±6.21Ab | 210.75±41.12Ab | 287.75±21.06Aa | 270.33±36.02Aab | 243.75±18.33Abc | 225.75±6.7Ac | 17.573*** | 0.396NS | 1.507NS |
| pH | 5.16±0.27Aa | 5.16±0.04Aa | 4.83±0.24Aab | 4.55±0.24Ab | 5.29±0.03Aa | 5.01±0.08Ab | 4.96±0.08Ab | 4.75±0.08Ac | 3.029* | 49.546*** | 0.551NS |
| UR (mg·g-1·24h-1) | 179.11±33.3Bd | 286.66±12.53Bc | 408.04±61.39Bb | 664.58±100.81Aa | 361.47±147Ab | 722.46±239.73Aab | 960.61±395.64Aab | 1062.5±640.64Aa | 6.223** | 14.885** | 0.578NS |
| NIR (mg·g-1·24h-1) | 9.42±5.24Aa | 9.40±5.61Aa | 5.96±3.88Aa | 4.60±2.78Aa | 8.34±2.63Aa | 11.10±2.43Aa | 8.90±3.19Aa | 6.05±3.54Aa | 2.427NS | 0.854NS | 0.388NS |
| INV (mg·g-1·24h-1) | 4.83±3.42Aa | 6.67±2.19Aa | 5.49±1.14Aa | 3.60±0.93Aa | 7.41±2.86Aa | 9.24±2.75Aa | 5.69±3.09Aa | 6.23±2.01Aa | 2.252NS | 5.258* | 0.474NS |
| PPO (mg·g-1·24h-1) | 8.23±4.84Aa | 6.35±0.49Aa | 6.38±1.19Aa | 4.92±0.75Ba | 6.96±1.53Ab | 11.1±5.04Ab | 6.64±1.19Ab | 20.34±10.37Aa | 2.805NS | 9.021** | 5.587** |
| CAT (mg·g-1·24h-1) | 4.79±2.54Aa | 2.12±1.04Ba | 2.95±1.20Ba | 4.31±5.53Aa | 13.71±6.93Aa | 12.67±1.02Aa | 12.65±3.74Aa | 14.52±13.60Aa | 0.23NS | 21.546*** | 0.028NS |
| ACP (mg·g-1·24h-1) | 19.64±3.16Bb | 25.09±0.84Bab | 32.78±12.03Aa | 23.05±2.53Bab | 33.34±4.54Ab | 37.67±3.33Ab | 38.65±4.87Ab | 49.40±5.08Aa | 5.395** | 56.23*** | 4.801** |
| 产量Yield (t·hm-2) | 1.11±0.35Bc | 2.60±0.51Bb | 4.90±1.10Aa | 5.88±0.63Aa | 3.60±0.25Ac | 4.69±0.23Ab | 5.31±0.68Aab | 6.03±0.53Aa | 34.331*** | 110.921*** | 5.258* |
表2
长期施氮量对茶园土壤真菌α多样性的影响"
| 季节 Seasons | 处理 Treatment | Sobs指数 Sobs index | Ace指数 Ace index | Chao1指数 Chao1 index | Shannon 指数 Shannon index | Simpson指数 Simpson index | 覆盖度 Coverage (%) |
|---|---|---|---|---|---|---|---|
| 春季 Spring | N0 | 822±45Aab | 906±51Aa | 905±48Aa | 4.31±0.21Aa | 0.04±0.01Bb | 99.82 |
| N1 | 858±146Aa | 969±135Aa | 974±141Aa | 4.16±0.61Aab | 0.07±0.05Aab | 99.77 | |
| N2 | 818±75Aab | 923±77Aa | 932±65Aa | 3.81±0.45Aab | 0.11±0.06Aab | 99.80 | |
| N3 | 676±69Ab | 745±70Ab | 745±69Ab | 3.51±0.29Ab | 0.12±0.03Aa | 99.85 | |
| 秋季 Autumn | N0 | 693±89Bab | 796±77Aab | 795±83Aab | 3.53±0.44Ba | 0.11±0.04Aa | 99.80 |
| N1 | 793±106Aa | 900±107Aa | 892±95Aa | 3.92±0.53Aa | 0.08±0.06Aa | 99.80 | |
| N2 | 702±12Bab | 838±37Aab | 839±32Ba | 3.49±0.22Aa | 0.12±0.04Aa | 99.78 | |
| N3 | 637±30Ab | 727±27Ab | 726±34Ab | 3.54±0.45Aa | 0.12±0.06Aa | 99.81 | |
| 施氮量Nitrogen (N) | 5.76** | 8.731** | 9.235** | 2.512NS | 1.884NS | ||
| 季节Seasons(S) | 9.089** | 6.174** | 7.48** | 4.586* | 1.88NS | ||
| 施氮量×季节N×S | 0.537NS | 0.466NS | 0.508NS | 1.226NS | 0.814NS | ||
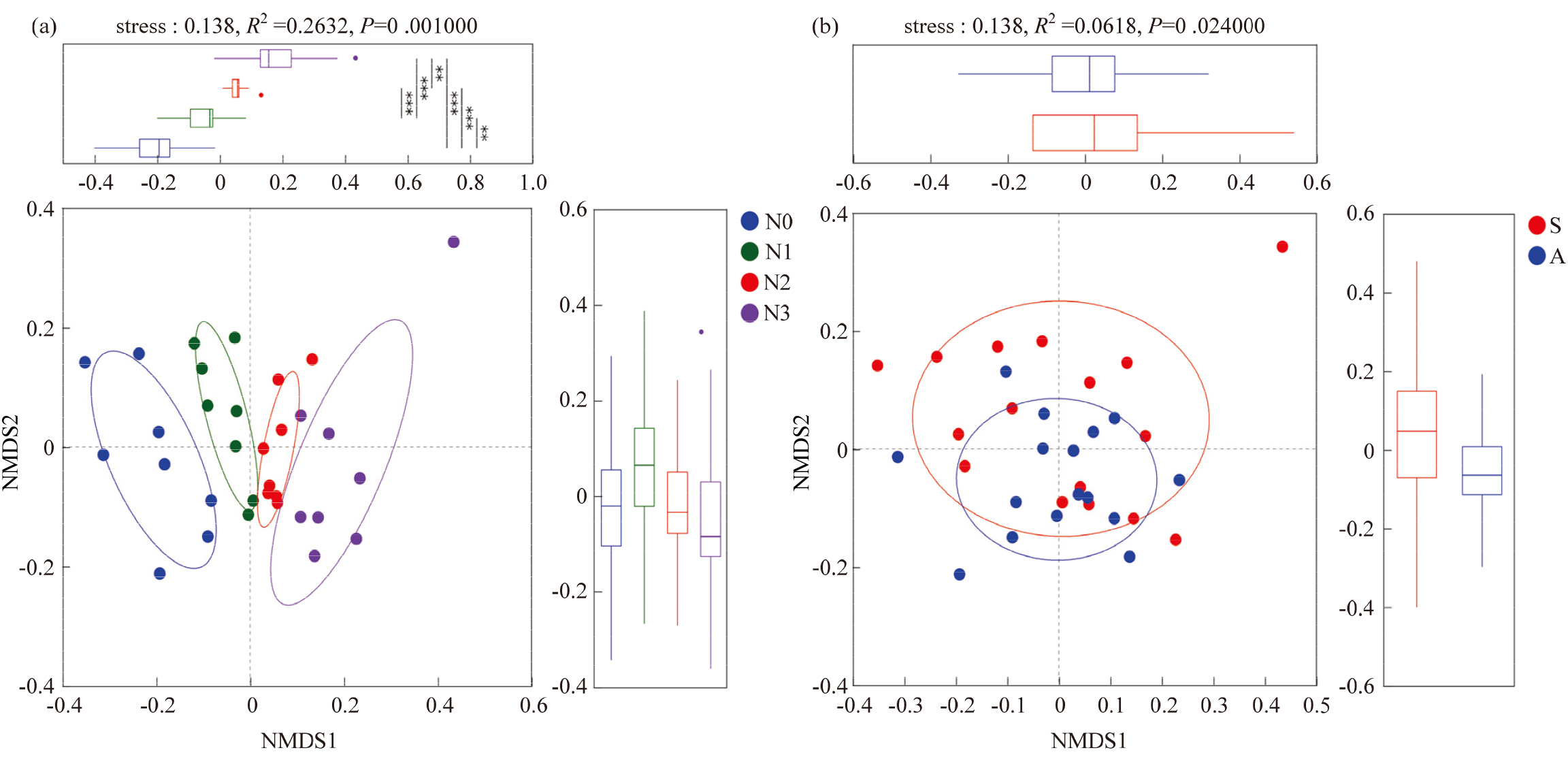
图2
基于Bray-Curtis 距离土壤真菌非度量多维尺度分析(NMDS)(a:不同施氮水平;b:不同采样季节;S:春季;A:秋季)"

图3
各样地土壤真菌群落组成在门水平(a)和属水平(b)上相对丰度(S:春季;A:秋季)"
表3
长期施氮和季节对茶园土壤真菌群落优势门和属相对丰度的双因素方差分析(F值)"
| 分类Classification | 因素Factor | 施氮量Nitrogen (N) | 季节Seasons(S) | 施氮量×季节 N×S |
|---|---|---|---|---|
| 门 Phylum | 子囊菌门Ascomycota | 0.448NS | 0.034NS | 0.382NS |
| 担子菌门Basidiomycota | 1.312NS | 2.267NS | 0.39NS | |
| 未知分类菌门 Unclassified_k__Fungi | 1.971NS | 1.999NS | 0.023NS | |
| 被孢霉门Mortierellomycota | 0.858NS | 9.239** | 1.066NS | |
| 罗兹菌门Rozellomycota | 4.998** | 1.516NS | 0.843NS | |
| 球囊菌门Glomeromycota | 3.198* | 2.062NS | 1.21NS | |
| 属 Genus | 沙蜥属Saitozyma | 1.287NS | 3.132NS | 0.512NS |
| 未知分类菌属 Unclassified_k__Fungi | 1.971NS | 1.999NS | 0.023NS | |
| 镰刀属Fusarium | 7.43** | 0.091NS | 0.504NS | |
| 木霉属 Trichoderma | 0.418NS | 2.213NS | 0.944NS | |
| 被孢霉属 Mortierella | 1.231NS | 8.847** | 0.992NS | |
| 青霉属Penicillium | 36.053** | 8.18** | 1.161NS | |
| 毛壳菌属 Chaetomium | 0.528NS | 7.6** | 0.889NS | |
| Unclassified_p__Ascomycota | 2.979* | 0.028NS | 0.421NS | |
| 枝孢菌属Cladosporium | 0.567NS | 3.435NS | 0.801NS | |
| Unclassified_c__Archaeorhizomycetes | 0.748NS | 0.843NS | 0.351NS |

图4
不同施氮处理土壤真菌的LEfSe分析(LDA阈值为3.5) A:春季;B:秋季(A: Spring; B: Autumn)"

图5
不同施氮量条件下茶园土壤真菌功能类型及非度量多维尺度分析(NMDS)分析 A:真菌功能类型;B:春季;C:秋季 A: Soil fungi function; B: Spring; C: Autumn"
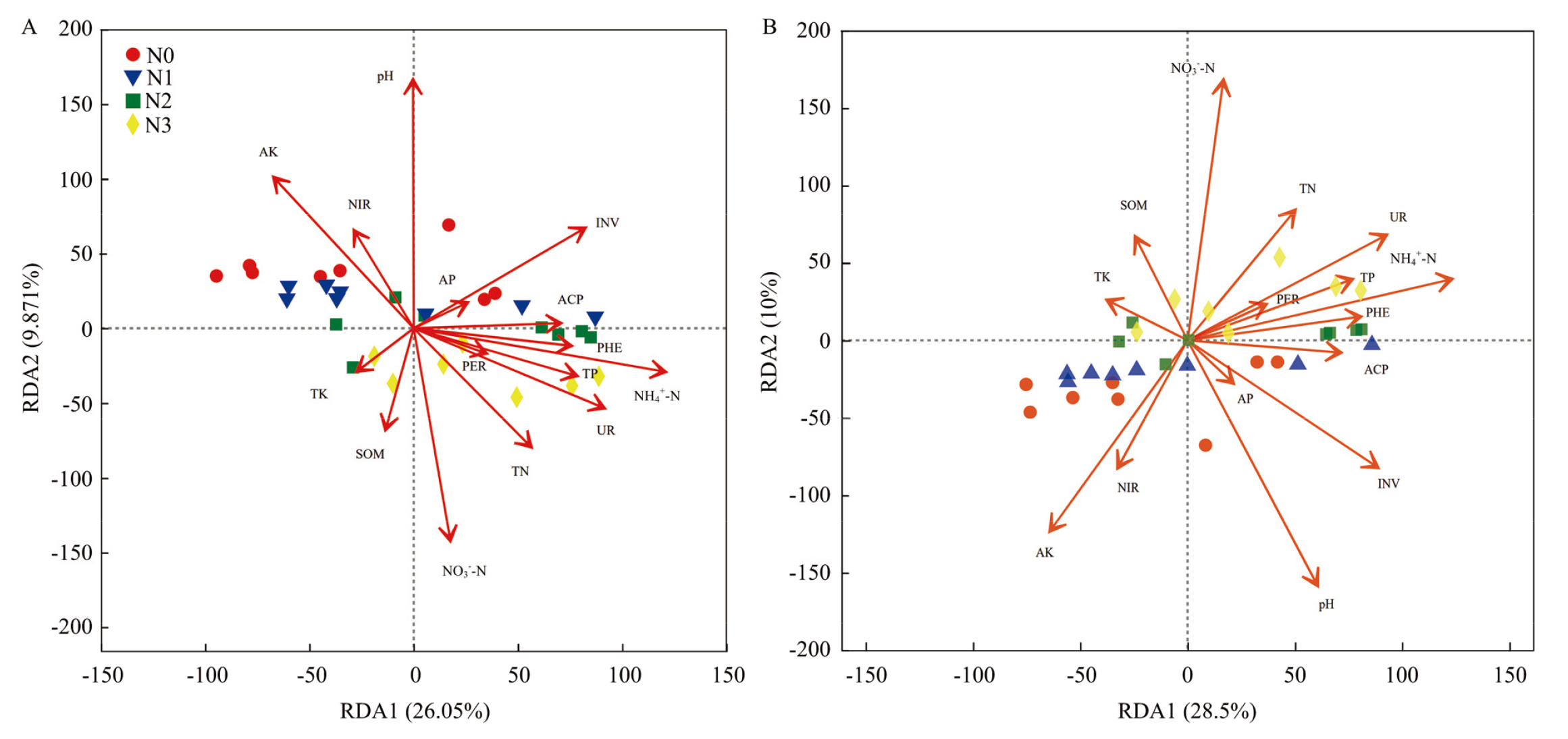
图6
土壤真菌群落结构(A)及功能类群(B)与土壤性质的冗余分析"

图7
不同施氮条件下土壤真菌分类学的共现性网络"
表4
茶园土壤真菌网络拓扑参数"
| 拓扑参数 Topological parameters | N0 | N1 | N2 | N3 |
|---|---|---|---|---|
| 边数Edges | 140 | 178 | 159 | 173 |
| 正相关边数Positive edges | 78 (55.71%) | 122 (68.54%) | 86 (54.09%) | 90 (52.02%) |
| 负相关边数Negative edges | 62 (44.29%) | 56 (31.46%) | 73 (45.91%) | 83 (47.98%) |
| 节点Nodes | 91 | 89 | 89 | 94 |
| 网络直径Network diameter | 22 | 11 | 18 | 12 |
| 模块性Modularity | 0.734 | 0.693 | 0.750 | 0.676 |
| 平均度Average degree | 3.077 | 4.000 | 3.573 | 3.681 |
| 平均路径长度Average path length | 7.364 | 4.871 | 6.889 | 5.188 |
| 平均聚类系数 Average clustering coefficient | 0.351 | 0.399 | 0.490 | 0.441 |
| 网络密度Network density | 0.034 | 0.045 | 0.041 | 0.040 |
| 模块数 Number of modules | 5 | 10 | 13 | 11 |
表5
不同施氮处理下茶园土壤真菌群落的构建"
| 季节 Seasons | 处理 Treatment | 确定性过程Determinism (%) | 随机性过程Stochasticity (%) | |||||
|---|---|---|---|---|---|---|---|---|
| 异质选择Heterogeneous selection | 同质选择Homogeneous selection | 比例之和 Total | 同质扩散 Homogeneous dispersal | 扩散限制 Dispersal limitation | 生态漂变 Drift | 比例之和 Total | ||
| 春季 Spring | N0 | 29.76 | 0.73 | 30.48 | 0.20 | 30.20 | 39.12 | 69.52 |
| N1 | 42.40 | 0.02 | 42.42 | 0.90 | 23.41 | 33.27 | 57.58 | |
| N2 | 41.10 | 0.16 | 41.26 | 0.24 | 23.50 | 34.99 | 58.74 | |
| N3 | 24.31 | 0.46 | 24.77 | 0.80 | 26.90 | 47.53 | 75.23 | |
| 秋季 Autumn | N0 | 36.90 | 0.01 | 36.91 | 1.23 | 21.28 | 40.58 | 63.09 |
| N1 | 48.96 | 0.10 | 49.06 | 0.13 | 20.67 | 30.14 | 50.94 | |
| N2 | 52.03 | 0.00 | 52.03 | 0.90 | 13.95 | 33.11 | 47.96 | |
| N3 | 51.08 | 0.37 | 51.45 | 1.17 | 11.57 | 35.81 | 48.55 | |
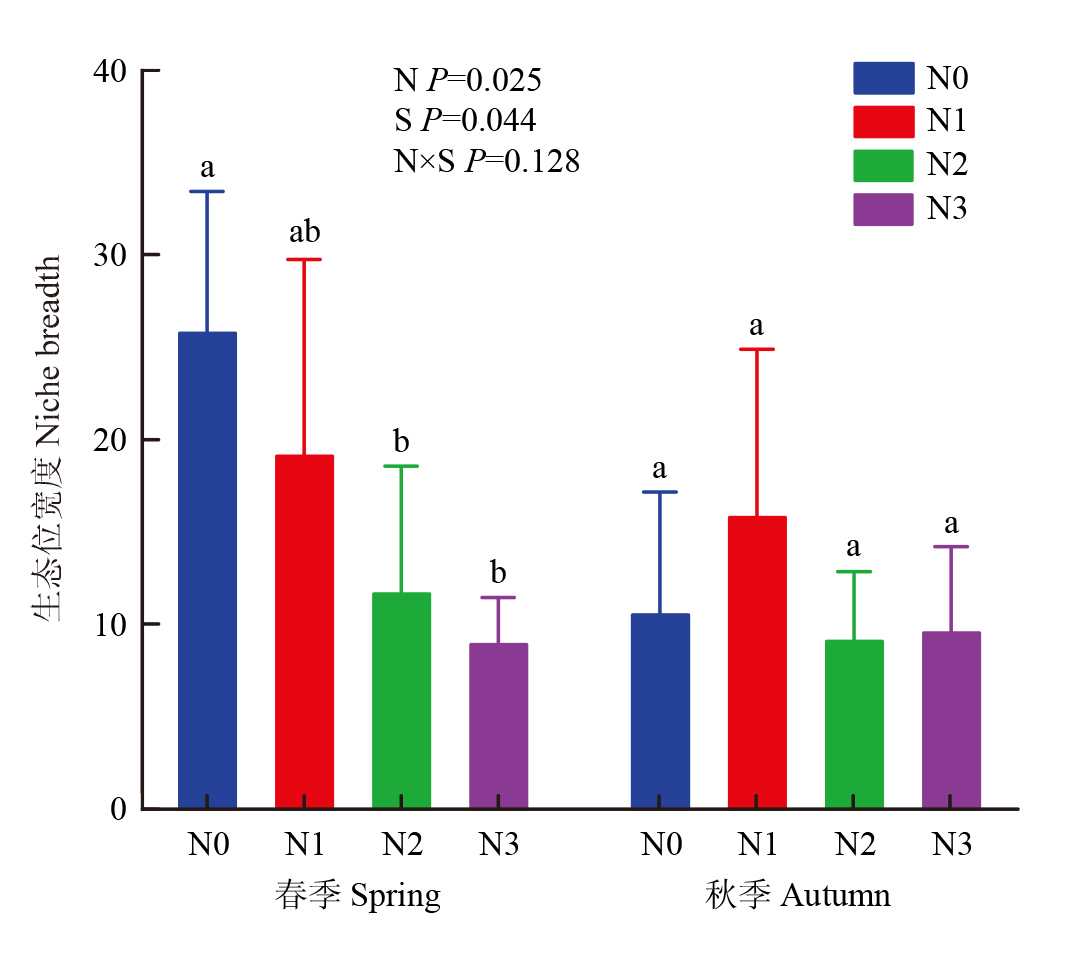
图8
不同施氮处理对真菌群落生态位宽度的影响"
| [1] |
doi: 10.1111/nph.v226.1 |
| [2] |
doi: 10.1007/s00248-022-02092-8 |
| [3] |
陈保冬, 付伟, 伍松林, 朱永官. 菌根真菌在陆地生态系统碳循环中的作用. 植物生态学报, 2024, 48(1): 1-20.
doi: 10.17521/cjpe.2023.0075 |
|
doi: 10.17521/cjpe.2023.0075 |
|
| [4] |
|
| [5] |
doi: 10.1038/s41558-021-00989-9 |
| [6] |
doi: S0981-9428(19)30066-X pmid: 30849677 |
| [7] |
doi: 10.3389/fpls.2023.1249202 |
| [8] |
doi: 10.1016/j.scitotenv.2019.136439 |
| [9] |
doi: 10.1007/s11356-018-2646-2 |
| [10] |
苏有健, 廖万有, 丁勇, 王宏树, 夏先江. 不同氮营养水平对茶叶产量和品质的影响. 植物营养与肥料学报, 2011, 17(6): 1430-1436.
|
|
|
|
| [11] |
doi: 10.1186/s12870-017-1111-6 |
| [12] |
王钊, 张冰, 董思奇, 胡雨翕, 齐书宇, 冯国忠, 高强, 周雪. 长期施用氮肥对黑土和风沙土根际微生物群落结构及功能的影响. 中国农业科学, 2025, 58(3): 520-536. doi: 10.3864/j.issn.0578-1752.2025.03.009.
|
|
|
|
| [13] |
doi: 10.1111/gcb.16746 pmid: 37186143 |
| [14] |
doi: 10.1111/gcb.2018.24.issue-8 |
| [15] |
doi: 10.1111/gcb.16588 pmid: 36607159 |
| [16] |
doi: 10.1128/spectrum.03003-22 |
| [17] |
doi: 10.1016/j.soilbio.2023.108982 |
| [18] |
季凌飞, 倪康, 马立锋, 陈兆杰, 赵远艳, 阮建云, 郭世伟. 不同施肥方式对酸性茶园土壤真菌群落的影响. 生态学报, 2018, 38(22): 8158-8166.
|
|
|
|
| [19] |
doi: 10.1007/s11104-022-05372-x |
| [20] |
doi: 10.1007/s11104-019-04291-8 |
| [21] |
doi: 10.3390/metabo11030146 |
| [22] |
doi: 10.1186/s40168-018-0570-9 pmid: 30336777 |
| [23] |
鲁如坤. 土壤农业化学分析方法. 北京: 中国农业科学技术出版社, 2000.
|
|
|
|
| [24] |
doi: 10.1016/j.funeco.2015.06.006 |
| [25] |
|
| [26] |
doi: 10.1038/s41467-020-18560-z pmid: 32948774 |
| [27] |
doi: 10.1016/j.scitotenv.2018.07.016 |
| [28] |
doi: 10.1016/j.apsoil.2021.103976 |
| [29] |
doi: 10.3390/foods13132091 |
| [30] |
doi: 10.1016/j.soilbio.2016.12.023 |
| [31] |
doi: 10.1088/1748-9326/10/2/024019 |
| [32] |
doi: 10.1016/j.soilbio.2014.11.004 |
| [33] |
doi: 10.1038/s41467-021-23605-y pmid: 34108462 |
| [34] |
doi: 10.1126/science.aau1361 pmid: 31604314 |
| [35] |
doi: 10.1111/ele.12754 pmid: 28295970 |
| [36] |
doi: 10.1046/j.1469-8137.2003.00714.x pmid: 33873396 |
| [37] |
doi: 10.1111/ele.13537 pmid: 32455502 |
| [38] |
doi: 10.1126/sciadv.aau4578 |
| [39] |
doi: 10.1093/jpe/rtaa099 |
| [40] |
靳海洋, 闫雅倩, 张德奇, 杨程, 岳俊芹, 李向东, 邵运辉, 方保停, 王汉芳, 秦峰. 秸秆还田下不同追氮量对麦田土壤真菌群落结构和生态网络的影响. 环境科学, 2023, 44(2): 1085-1094.
|
|
doi: 10.1021/es9019645 |
|
| [41] |
doi: 10.1007/s11368-020-02629-9 |
| [42] |
doi: 10.1038/s41467-019-11787-5 pmid: 31444343 |
| [43] |
doi: 10.1016/j.soilbio.2020.108119 |
| [44] |
刘蕾, 肖广敏, 王凌, 茹淑华, 张国印, 孙世友. 设施蔬菜土壤丛枝菌根真菌多样性及群落构建的季相变化. 土壤学报, 2024, 61(1): 200-210.
|
|
|
|
| [45] |
doi: 10.1002/imt2.106 pmid: 38868425 |
| [46] |
doi: 10.1016/j.isci.2022.103893 |
| [47] |
doi: 10.1128/msystems.01248-24 |
| [48] |
doi: 10.1002/imt2.18 pmid: 38868564 |
| [49] |
doi: 10.1016/j.soilbio.2020.108023 |
| [50] |
doi: 10.1016/j.scitotenv.2024.172100 |
| [51] |
doi: 10.1111/gcb.v27.16 |
| [52] |
doi: 10.1016/j.soilbio.2015.07.007 |
| [1] | 王仁卓, 李玥颖, 黄绍敏, 姜桂英, 张琦, 刘超麟, 杨锦, 王梦茹, 王贝贝, 刘芳, 郭斗斗, 介晓磊, 宋练, 刘世亮. 长期有机无机肥配施对潮土细菌群落结构、生态网络及关键物种的影响[J]. 中国农业科学, 2026, 59(2): 354-367. |
| [2] | 潘丽媛, 王永军, 李海军, 侯富, 李菁, 李丽丽, 孙苏阳. 基于转录组和WGCNA筛选小麦籽粒蛋白质累积相关调控基因[J]. 中国农业科学, 2025, 58(6): 1065-1082. |
| [3] | 穆赢通, 路景诗, 张雨桐, 石凤翎. 基于转录组和WGCNA的直立型花苜蓿抗旱关键基因识别[J]. 中国农业科学, 2025, 58(21): 4528-4543. |
| [4] | 董雪, 陈梦秋, 邵晋, 吴学友, 唐培安. 基于WGCNA的稻谷储藏期间差异基因挖掘与品质调控网络构建[J]. 中国农业科学, 2025, 58(14): 2885-2903. |
| [5] | 宣泽鹏, 冯慧瑶, 陈美淇, 徐基胜, 刘梦璇, 赵炳梓, 张佳宝. 秸秆还田配施化肥对土壤生态系统多功能性的影响[J]. 中国农业科学, 2025, 58(14): 2821-2837. |
| [6] | 栗海鹏, 杜武焰, 吴涵茜, 张杰, 孟会生, 洪坚平, 徐明岗, 郝鲜俊, 高文俊. 不同有机肥对煤矿复垦区土壤养分及细菌群落结构的影响[J]. 中国农业科学, 2024, 57(16): 3207-3219. |
| [7] | 陈晓涓, 王海菊, 王富敏, 雍清青, 黄顺满, 屈燕. 基于WGCNA鉴定全缘叶绿绒蒿类黄酮合成途径关键基因[J]. 中国农业科学, 2024, 57(15): 3053-3070. |
| [8] | 彭佳伟, 张叶, 寇单单, 杨丽, 刘晓飞, 张学英, 陈海江, 田义. ‘仓方早生’桃及其早熟芽变不同发育时期果实的转录组分析[J]. 中国农业科学, 2023, 56(5): 964-980. |
| [9] | 郭晗玥, 王东升, 阮杨, 乔亦铸, 张芸滔, 李玲, 黄启为, 郭世伟, 凌宁, 沈其荣. 西瓜连作根际土壤微生物群落演替特征[J]. 中国农业科学, 2023, 56(21): 4245-4258. |
| [10] | 刘蕾, 史建硕, 张国印, 郜静, 李玭, 任燕利, 王丽英. 长期施有机肥对设施番茄土壤稀有和丰富细菌亚群落的影响[J]. 中国农业科学, 2023, 56(18): 3615-3628. |
| [11] | 张杰,王川,董晓霞,祝文琪,岳慧丽,刘升平,周清波. 基于裂变模式的农业农村信息快速调查及分析平台开发与应用[J]. 中国农业科学, 2022, 55(21): 4158-4174. |
| [12] | 赵卫松,郭庆港,苏振贺,王培培,董丽红,胡卿,鹿秀云,张晓云,李社增,马平. 马铃薯健株与黄萎病株根际土壤真菌群落结构及其对碳源利用特征[J]. 中国农业科学, 2021, 54(2): 296-309. |
| [13] | 黄子粤,刘文君,覃仁柳,庞师婵,肖健,杨尚东. 不同品种南瓜内生细菌多样性及PICRUSt基因功能预测分析[J]. 中国农业科学, 2021, 54(18): 4018-4032. |
| [14] | 颜鹏,韩文炎,李鑫,张丽平,张兰. 中国茶园土壤酸化现状与分析[J]. 中国农业科学, 2020, 53(4): 795-801. |
| [15] | 李文广,杨晓晓,黄春国,薛乃雯,夏清,刘小丽,张晓琪,杨思,杨珍平,高志强. 饲料油菜作绿肥对后茬麦田土壤肥力及细菌群落的影响[J]. 中国农业科学, 2019, 52(15): 2664-2677. |
|
||