






中国农业科学 ›› 2026, Vol. 59 ›› Issue (6): 1286-1301.doi: 10.3864/j.issn.0578-1752.2026.06.011
曹海顺1( ), 周东源1,2(), 王瑞1, 施招婉1, 吴廷全1, 张长远1,2()
), 周东源1,2(), 王瑞1, 施招婉1, 吴廷全1, 张长远1,2()
收稿日期:2025-10-16
接受日期:2025-12-05
出版日期:2026-03-24
发布日期:2026-03-24
通信作者:
联系方式:
曹海顺,E-mail:caohaishun@gdaas.cn。周东源,E-mail:804698010@qq.com。曹海顺和周东源为同等贡献作者。
基金资助:
CAO HaiShun1(), ZHOU DongYuan1,2(), WANG Rui1, SHI ZhaoWan1, WU TingQuan1, ZHANG ChangYuan1,2()
Received:2025-10-16
Accepted:2025-12-05
Published:2026-03-24
Online:2026-03-24
摘要:
【目的】黄瓜是我国设施栽培第一大蔬菜。然而,频发的设施弱光胁迫易造成黄瓜苗过度徒长。筛选弱光胁迫不易徒长种质,并挖掘相关基因,为黄瓜耐弱光性状精准遗传改良提供参考。【方法】利用弱光胁迫评价体系筛选黄瓜种质,构建分离群体,采用BSA-seq分析性状关联位点,综合家族基因和功能分析挖掘候选基因。【结果】弱光胁迫下获得2份下胚轴长度差异材料CS(长下胚轴)和CR(短下胚轴)。经不同光质处理,发现CS和CR对远红光具有响应差异。以CS和CR为亲本,构建F1群体和F2群体。以弱光胁迫下胚轴长度为性状参数,结合BSA-seq的方法在黄瓜中定位到2个显著与下胚轴长度相关的新位点:LSH1(Chr.4)和LSH2(Chr.5),位点内分别包含371和163个基因。进一步对远红光信号通路的关键基因PHYA、FHL和FRS类转录因子进行全面鉴定,共获得2个远红光受体基因CsPHYA1和CsPHYA2,皆位于第6染色体;1个远红光受体转运基因CsFHY1,位于第3染色体;22个FRS转录因子,除第2染色体外,其他染色体均有分布。其中,CsFRS12位于第4染色体LSH1位点内,进化树和保守结构域分析显示,CsFRS12具有保守的FAR1 DNA结合结构域、MULE转座酶结构域和SWIM锌指蛋白结构域,与远红光信号关键转录因子AtFRS2、AtFAR1和AtFHY3关系较近,同属于Ⅰ亚家族成员。经克隆,CsFRS12CR和CsFRS12CS编码序列中存在多个单核苷酸突变(SNP),与下胚轴长度显著相关。进一步通过酵母转录活性分析,发现CsFRS12CR转录活性低于CsFRS12CS。CsFRS12在植株茎和幼苗下胚轴中表达量较高,即CsFRS12是调控弱光胁迫黄瓜下胚轴伸长的关键候选基因。【结论】获得2份弱光胁迫响应差异黄瓜材料,定位到2个调控弱光胁迫响应的新位点LSH1和LSH2,在LSH1位点中候选到1个远红光信号调控的关键转录因子CsFRS12,可能在调控黄瓜适应弱光胁迫中发挥关键作用。
曹海顺, 周东源, 王瑞, 施招婉, 吴廷全, 张长远. 弱光胁迫短下胚轴黄瓜种质鉴定及其遗传位点挖掘[J]. 中国农业科学, 2026, 59(6): 1286-1301.
CAO HaiShun, ZHOU DongYuan, WANG Rui, SHI ZhaoWan, WU TingQuan, ZHANG ChangYuan. Identification of Short Hypocotyl Cucumber Germplasm Under Low Light Stress and QTL Mapping of the Trait[J]. Scientia Agricultura Sinica, 2026, 59(6): 1286-1301.
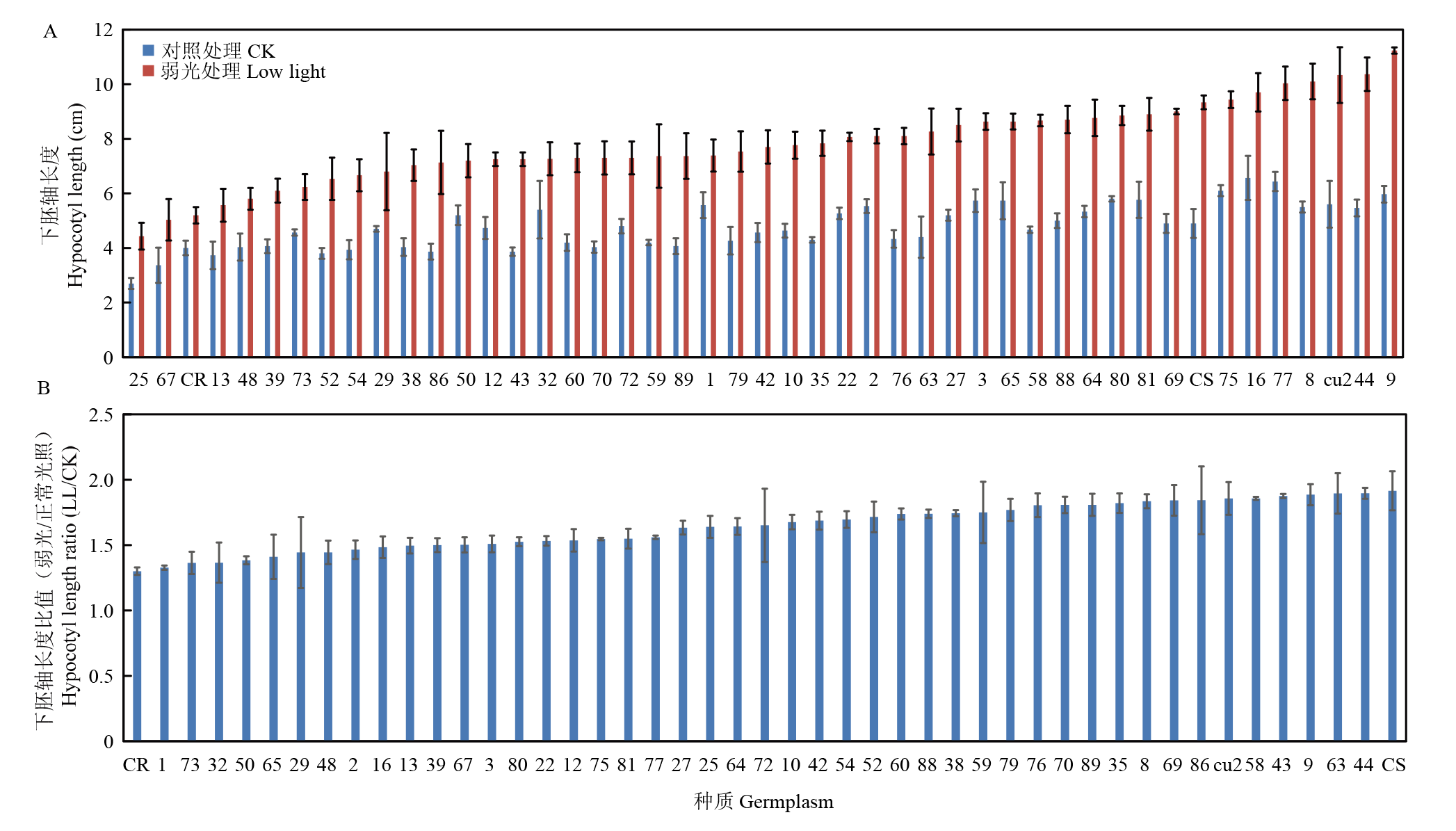
图1
黄瓜下胚轴对弱光胁迫的敏感度分析 A:不同黄瓜种质在弱光胁迫和正常光照下的下胚轴长度值;B:弱光胁迫和正常光照下不同黄瓜种质下胚轴长度比值"

图2
弱光胁迫下黄瓜下胚轴长度的表型以及性状的定位与分析 A:弱光胁迫下CS、CR和F1的表型;B:弱光胁迫下CS、CR和F1的下胚轴长度值;C:黄瓜F2群体下胚轴长度数值分布;D:Δ(SNP指数)在黄瓜染色体上分布的曼哈顿图。红线表示关联阈值99%,蓝线表示关联阈值95%"
表1
弱光胁迫下黄瓜下胚轴长度QTL的BSA-seq定位结果"
| 方法 Method | 数量性状基因座名称 QTL name | 起始位点 Start position (bp) | 终止位点 End position (bp) | 染色体 Chromosome | 峰值 Peak | 基因数目 Gene numbers |
|---|---|---|---|---|---|---|
| 单核苷酸多态性指数 Δ(SNP-index) | LSH1 | 20300001 | 24200000 | Chr.4 | 0.505331835 | 371 |
| LSH2 | 6700001 | 10200000 | Chr.5 | 0.532489898 | 163 | |
| G统计值 G-statistic | LSH1 | 21100001 | 23300000 | Chr.4 | 11.26343258 | 196 |
| LSH2 | 7100001 | 9200000 | Chr.5 | 13.22583847 | 74 | |
| 欧氏距离 Euclidean distance | LSH1 | 21000001 | 23500000 | Chr.4 | 0.489802113 | 229 |
| LSH2 | 8000001 | 9200000 | Chr.5 | 0.463019996 | 37 |
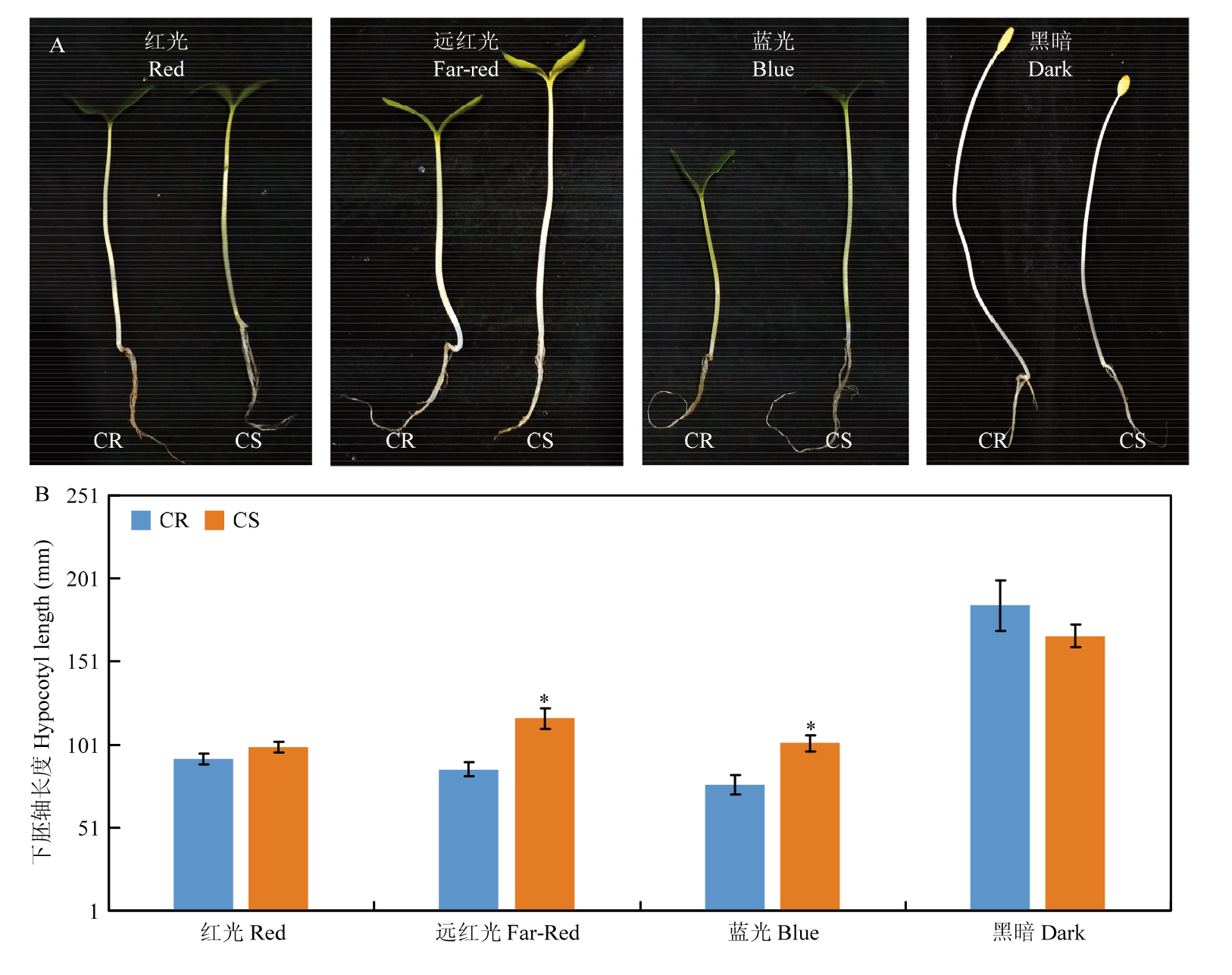
图3
不同光质(红光、远红光、蓝光和黑暗)处理下黄瓜下胚轴的表型(A)和长度(B)"
表2
黄瓜FRS家族基因基本信息"
| 基因序号 Gene ID | 基因名称 Name | 染色体 Chromosome | 起始位点 Start position (bp) | 终止位点 Stop position (bp) | 正反链 Strand | 编码框长度 CDS length (bp) | 蛋白长度 Protein length (aa) |
|---|---|---|---|---|---|---|---|
| CsaV3_1G010190.1 | CsFRS1 | Chr.1 | 6322731 | 6327319 | - | 2565 | 855 |
| CsaV3_1G042070.1 | CsFRS2 | Chr.1 | 26879471 | 26883820 | + | 2628 | 876 |
| CsaV3_3G013860.1 | CsFRS3 | Chr.3 | 10392246 | 10397538 | - | 2364 | 788 |
| CsaV3_3G034800.1 | CsFHY1 | Chr.3 | 29369257 | 29371197 | + | 1041 | 347 |
| CsaV3_3G039150.1 | CsFRS4 | Chr.3 | 32174695 | 32179211 | + | 1170 | 390 |
| CsaV3_3G046280.1 | CsFRS5 | Chr.3 | 37806932 | 37811364 | + | 1818 | 606 |
| CsaV3_4G001920.1 | CsFRS6 | Chr.4 | 1130443 | 1133692 | + | 2232 | 744 |
| CsaV3_4G001930.1 | CsFRS7 | Chr.4 | 1135597 | 1138623 | + | 2007 | 669 |
| CsaV3_4G003380.1 | CsFRS8 | Chr.4 | 2056681 | 2059569 | + | 765 | 255 |
| CsaV3_4G003760.1 | CsFRS9 | Chr.4 | 2290171 | 2295355 | + | 750 | 250 |
| CsaV3_4G007310.1 | CsFRS10 | Chr.4 | 4973487 | 4978240 | - | 2424 | 808 |
| CsaV3_4G007320.1 | CsFRS11 | Chr.4 | 4978958 | 4986126 | - | 2538 | 846 |
| CsaV3_4G032340.1 | CsFRS12 | Chr4 | 22862562 | 22866893 | + | 2325 | 775 |
| CsaV3_5G014020.1 | CsFRS13 | Chr.5 | 10880232 | 10889015 | - | 2040 | 680 |
| CsaV3_6G009980.1 | CsFRS14 | Chr.6 | 8087498 | 8091321 | + | 1488 | 496 |
| CsaV3_6G036060.1 | CsPHYA1 | Chr.6 | 20015890 | 20021182 | - | 3450 | 1150 |
| CsaV3_6G036100.1 | CsPHYA2 | Chr.6 | 20046075 | 20053349 | + | 3375 | 1125 |
| CsaV3_6G048310.1 | CsFRS15 | Chr.6 | 28368052 | 28370312 | + | 2076 | 692 |
| CsaV3_6G053050.1 | CsFRS16 | Chr.6 | 30971693 | 30975462 | - | 2241 | 747 |
| CsaV3_7G002210.1 | CsFRS17 | Chr.7 | 1754310 | 1760052 | + | 2370 | 790 |
| CsaV3_7G002550.1 | CsFRS18 | Chr.7 | 1993692 | 2004852 | + | 2316 | 772 |
| CsaV3_7G026930.1 | CsFRS19 | Chr.7 | 16558935 | 16562421 | - | 666 | 222 |
| CsaV3_7G027310.1 | CsFRS20 | Chr.7 | 16862778 | 16871926 | + | 1491 | 497 |
| CsaV3_7G032290.1 | CsFRS21 | Chr.7 | 20436138 | 20438129 | + | 1989 | 663 |
| CsaV3_UNG168310.1 | CsFRS22 | Scaffold.77 | 105440 | 114205 | + | 2028 | 676 |
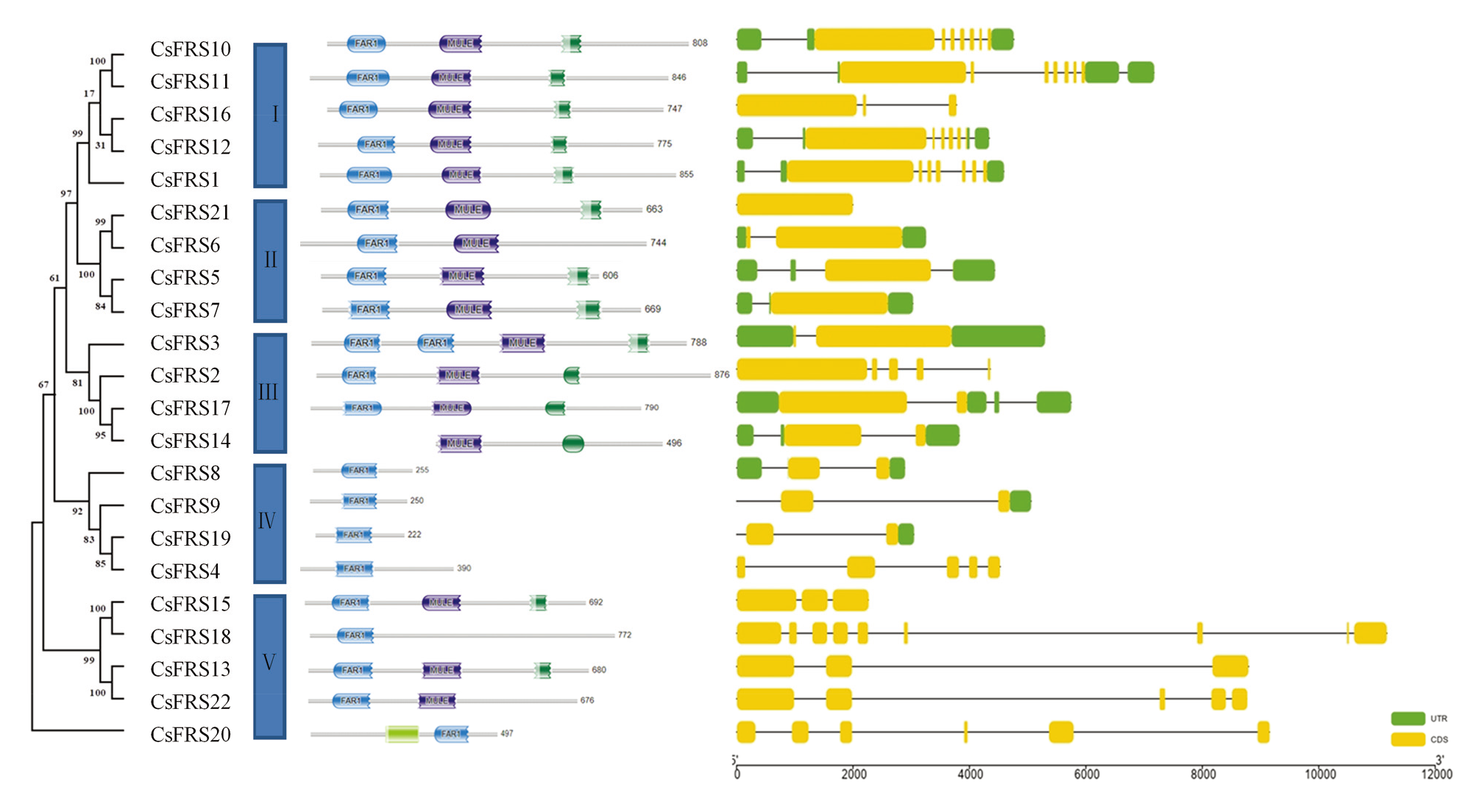
图4
黄瓜FRS家族蛋白的保守结构域和基因结构分析"
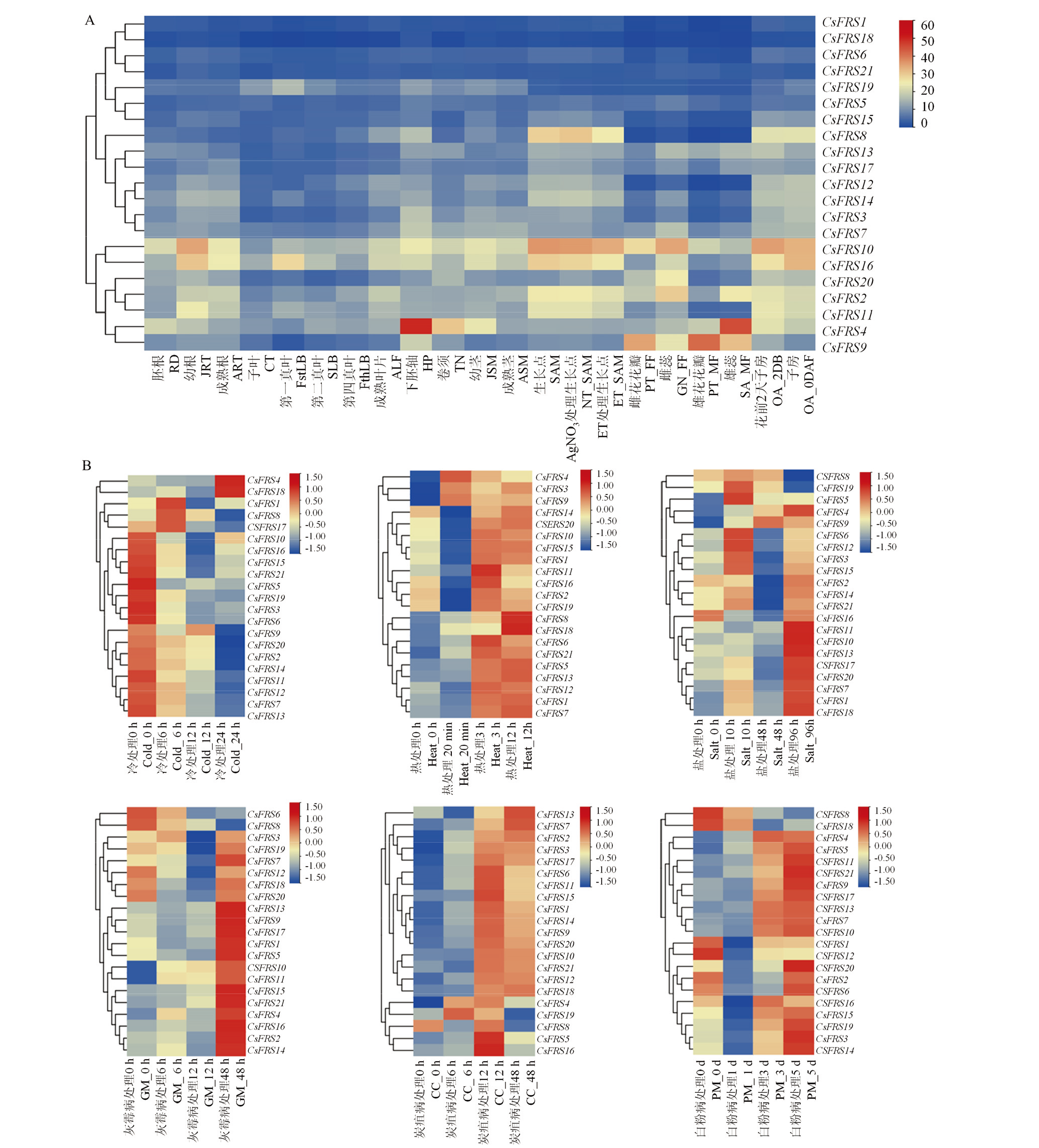
图5
黄瓜CsFRS家族基因的表达分析 A:组织表达分析;B:生物和非生物胁迫的诱导表达模式分析。HP:下胚轴;RD:胚根;CT:子叶;FstLB:第一真叶;SLB:第二真叶;FthLB:第四真叶;JSM:幼茎;JRT:幼根;SAM:生长点;ART:成熟根系;ALF:成熟叶片;ASM:成熟茎;TN:卷须;OA_2DBF:开花前两天子房;OA_0DAF:子房;PT_MF:雄花花瓣;SA_MF:雄蕊;PT_FF:雌花花瓣;GN_FF:雌蕊;EN:果实内果皮;MS:果实中果皮;EX:果实外果皮;NC:果把;SD:种子"

图6
黄瓜CsFRS家族基因的染色体分布、共线性和进化树分析 A:CsFRS家族基因的染色体分布和共线性分析;B:CsFRS家族基因的进化树分析。AT:拟南芥;Cla:西瓜;Csa:黄瓜;Cmo:南瓜;MELO:甜瓜;Solyc:番茄;Moc:苦瓜"

图7
黄瓜CR和CS中CsFRS12核苷酸和氨基酸序列的差异分析"

图8
黄瓜CsFRS12的转录活性和组织特异性表达分析 A:转录活性分析;B:CsFRS12表达分析"
| [1] |
doi: 10.1093/plcell/koaa018 |
| [2] |
|
| [3] |
doi: 10.1093/plphys/kiae153 pmid: 38478469 |
| [4] |
doi: 10.3389/fpls.2022.1093859 |
| [5] |
doi: 10.1146/annurev-arplant-050213-040145 pmid: 24471835 |
| [6] |
doi: 10.1104/pp.112.193359 |
| [7] |
doi: S0378-1119(19)30428-7 pmid: 31034942 |
| [8] |
doi: 10.1146/annurev-arplant-043015-112252 pmid: 26905653 |
| [9] |
doi: 10.1016/j.conb.2015.01.013 pmid: 25638281 |
| [10] |
doi: 10.1093/pcp/pcy153 |
| [11] |
doi: 10.15252/embr.201845762 |
| [12] |
doi: 10.1080/09168451.2015.1065171 |
| [13] |
doi: 10.1038/ncomms5636 |
| [14] |
doi: 10.1104/pp.110.162057 |
| [15] |
doi: 10.1104/pp.20.00587 pmid: 33889988 |
| [16] |
doi: 10.1038/s41467-020-15893-7 |
| [17] |
doi: S0960-9822(16)31257-X pmid: 27889265 |
| [18] |
doi: 10.1007/s00239-004-0294-2 pmid: 16170454 |
| [19] |
doi: 10.1093/plcell/koab302 |
| [20] |
doi: 10.1016/S0044-328X(80)80208-X |
| [21] |
|
| [22] |
doi: 10.1186/s43897-021-00005-w |
| [23] |
doi: 10.1093/jxb/erad181 |
| [24] |
doi: 10.1007/s00122-021-03849-4 |
| [25] |
doi: 10.3389/fpls.2024.1371435 |
| [26] |
|
| [27] |
doi: S1674-2052(20)30187-8 pmid: 32585190 |
| [28] |
doi: 10.1016/j.molp.2023.09.010 |
| [29] |
|
| [30] |
|
| [31] |
doi: 10.1199/tab.0148 |
| [32] |
|
| [33] |
doi: 10.1146/annurev-arplant-050312-120221 pmid: 23373700 |
| [34] |
doi: 10.1016/j.molp.2020.11.016 pmid: 33249237 |
| [35] |
doi: 10.1093/pcp/pcz191 pmid: 31617558 |
| [36] |
doi: 10.1146/arplant.1982.33.issue-1 |
| [37] |
doi: 10.3389/fpls.2019.00322 pmid: 30984211 |
| [38] |
doi: 10.3389/fpls.2022.857616 |
| [39] |
doi: 10.1111/pce.v47.2 |
| [40] |
doi: 10.3389/fpls.2024.1340260 |
| [41] |
doi: 10.1371/journal.pgen.1000143 |
| [42] |
doi: 10.1016/j.xplc.2019.100007 |
| [43] |
doi: 10.1126/science.1146281 |
| [44] |
doi: 10.3389/fpls.2016.00175 pmid: 26941752 |
| [45] |
doi: 10.1016/j.tplants.2015.04.003 |
| [46] |
doi: 10.1093/emboj/21.6.1339 |
| [47] |
doi: 10.1101/gad.13.15.2017 |
| [1] | 张利东, 郭一聪, 黄洪宇, 聂静, 王兵, 李梦雨, 李加旺, 眭晓蕾, 李愚鹤. 基于感官评价结合营养品质特征与风味物质特性关联分析黄瓜果实品质[J]. 中国农业科学, 2026, 59(5): 1087-1100. |
| [2] | 付涵, 于杨, 艾妞, 张思晴, 于连伟, 孙书豪, 赵金章, 韩晓玉, 施艳, 杨雪. 光合作用系统Ⅱ相关蛋白NbPsbQ1通过促进光合效率抑制病毒侵染[J]. 中国农业科学, 2026, 59(1): 90-100. |
| [3] | 刘宏祥, 张雪萍, 王逸飞, 王志成, 顾昊天, 宋卫涛, 陶志云, 徐文娟, 章双杰, 卢立志, 李慧芳, 朱春红. 金定鸭产蛋数性状的全基因组关联研究[J]. 中国农业科学, 2025, 58(15): 3145-3158. |
| [4] | 廖锴, 李欣, 石延霞, 谢学文, 李磊, 范腾飞, 王绍辉, 李宝聚, 柴阿丽. 塑料拱棚不同通风方式对黄瓜细菌性角斑病传播的影响[J]. 中国农业科学, 2025, 58(10): 1934-1946. |
| [5] | 姜兴林, 于连伟, 付涵, 艾妞, 崔荧钧, 李好海, 夏子豪, 袁虹霞, 李洪连, 杨雪, 施艳. 转录因子NbMYB1R1通过促进活性氧积累抑制病毒侵染[J]. 中国农业科学, 2024, 57(8): 1490-1505. |
| [6] | 赵猛, 毕焕改, 孟令浩, 姜婷婷, 张晓伟, 艾希珍. 低温循环诱导的胁迫记忆调控黄瓜耐冷性机制[J]. 中国农业科学, 2024, 57(24): 4933-4944. |
| [7] | 张开京, 何帅帅, 贾利, 胡玉超, 杨德坤, 陆晓民, 张其安, 严从生. 黄瓜DIR家族基因的全基因组鉴定及其表达分析[J]. 中国农业科学, 2023, 56(4): 711-728. |
| [8] | 朱莜芸, 曾玉玲, 李博, 袁玉洁, 周星, 李秋萍, 何辰延, 陈勇, 王丽, 程红, 周伟, 陶有凤, 雷小龙, 任万军, 邓飞. 花后弱光胁迫对成都平原籼稻米饭食味品质的影响[J]. 中国农业科学, 2023, 56(3): 430-440. |
| [9] | 王壮壮, 董邵云, 周琪, 苗晗, 刘小萍, 徐奎鹏, 顾兴芳, 张圣平. 黄瓜果实维生素C合成关键基因克隆与分析[J]. 中国农业科学, 2023, 56(3): 508-518. |
| [10] | 孙艳发, 吴琼, 林如龙, 陈红萍, 甘秋云, 沈玥, 王亚茹, 薛鹏飞, 陈飞帆, 刘健涛, 周陈鑫, 兰诗诗, 潘浩哲, 邓凡, 岳稳, 江宵兵, 李焰. 龙岩山麻鸭蛋品质性状的全基因组关联研究[J]. 中国农业科学, 2023, 56(3): 572-586. |
| [11] | 林萍, 王开良, 姚小华, 任华东. 基于转录组SNP构建油茶主要品种资源的分子身份证[J]. 中国农业科学, 2023, 56(2): 217-235. |
| [12] | 柴阿丽, 杨红敏, 王少骅, 赵昆, 高苇, 石延霞, 谢学文, 李磊, 范腾飞, 李宝聚. 湿度调控设施黄瓜棒孢叶斑病菌产孢和释放规律及防治技术[J]. 中国农业科学, 2023, 56(15): 2907-2918. |
| [13] | 于连伟, 姜兴林, 杨灵玲, 王贺, 张玉阳, 谢莉娜, 夏子豪, 李洪连, 杨雪, 施艳. 转录因子NbERF RAP2-1在黄瓜绿斑驳花叶病毒侵染中的功能[J]. 中国农业科学, 2023, 56(15): 2919-2928. |
| [14] | 冯向君, 王宏宇, 于静, 池春玉, 丁国华. 过表达拟南芥NPR1增强黄瓜对枯萎病和白粉病的抗性[J]. 中国农业科学, 2023, 56(14): 2701-2712. |
| [15] | 梁海文, 兰平秀, 柳勤海, 谭冠林, 陈小姣, 赵雁, 李凡. 侵染花叶青木的病毒种类鉴定及其基因序列分析[J]. 中国农业科学, 2023, 56(10): 1893-1904. |
|
||