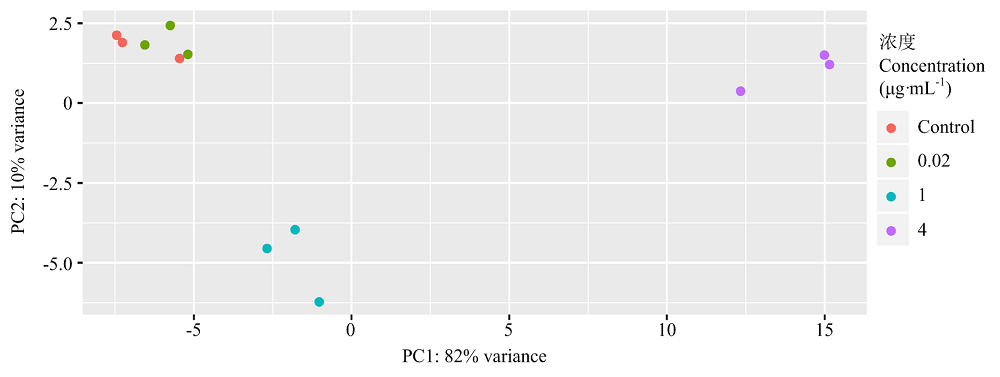
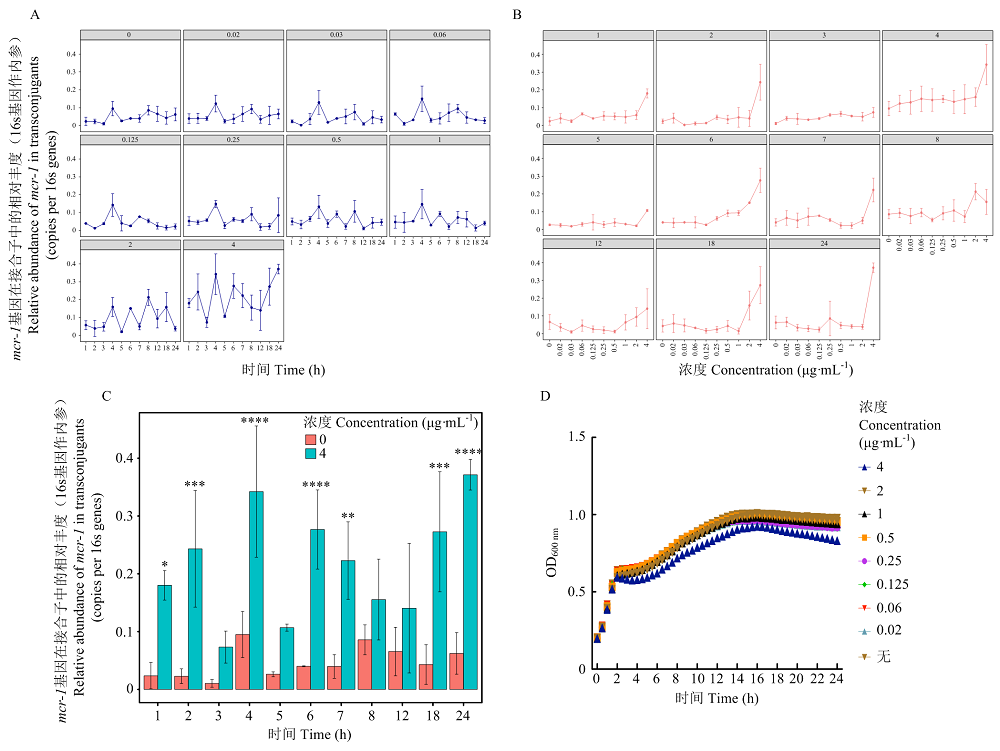
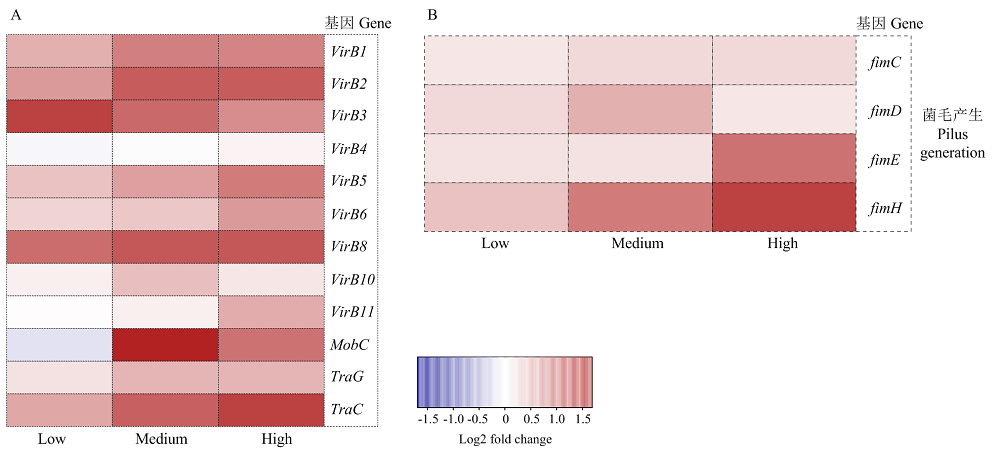
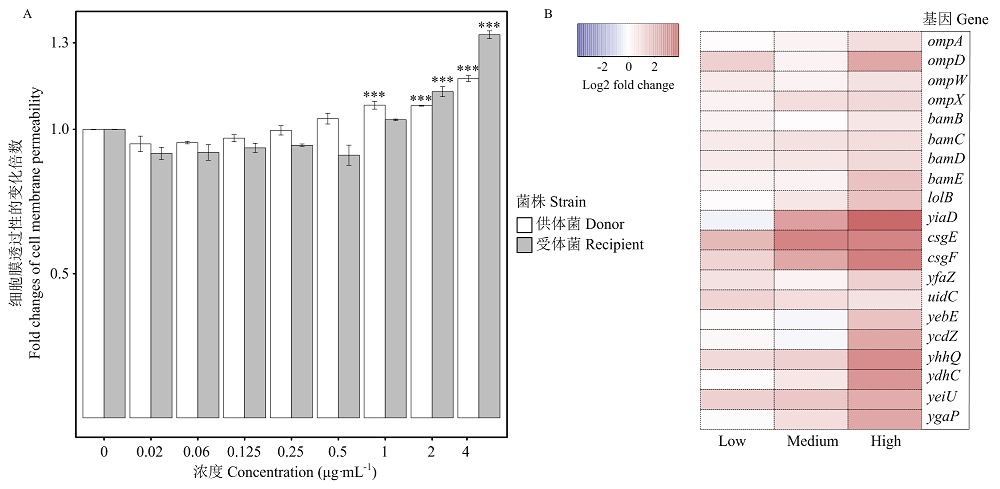
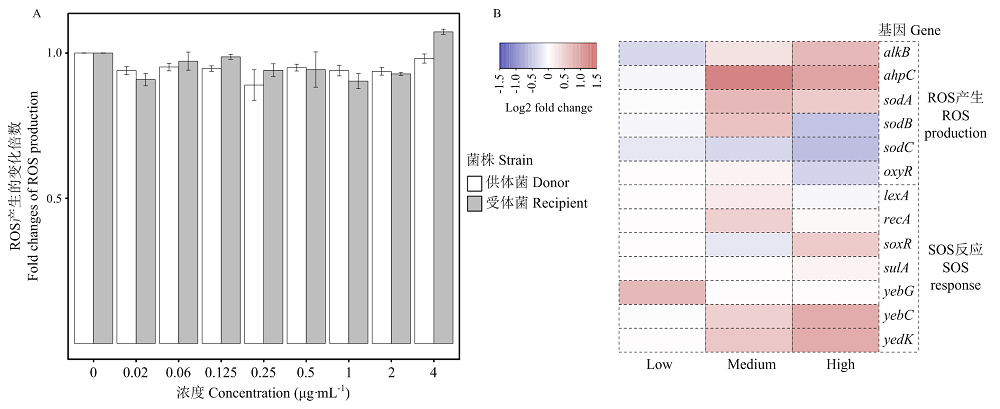
Scientia Agricultura Sinica ›› 2022, Vol. 55 ›› Issue (14): 2862-2874.doi: 10.3864/j.issn.0578-1752.2022.14.015
• ANIMAL SCIENCE·VETERINARY SCIENCE • Previous Articles
WANG XueYang1( ),JIANG JunYao1,YANG Lu1,SHAO DongYan1,WU CongMing1,SHEN JianZhong1,SHEN YingBo1,2(),WANG Yang1()
),JIANG JunYao1,YANG Lu1,SHAO DongYan1,WU CongMing1,SHEN JianZhong1,SHEN YingBo1,2(),WANG Yang1()
| [1] | 郝海红, 程古月, 戴梦红, 王旭, 王玉莲, 黄玲利, 刘振利, 袁宗辉. 对动物饲料中禁用抗菌促生长剂的反思. 中国农业科学, 2015, 48(3): 594-603. |
| HAO H H, CHENG G Y, DAI M H, WANG X, WANG Y L, HUANG L L, LIU Z L, YUAN Z H. Rethinking the Withdrawal of Antimicrobial Growth Promotants in Animal Feed. Scientia Agricultura Sinica, 2015, 48(3): 594-603. (in Chinese) | |
| [2] |
SHEN Y B, ZHANG R, SCHWARZ S, WU C M, SHEN J Z, WALSH TR, WANG Y. Farm animals and aquaculture: significant reservoirs of mobile colistin resistance genes. Environmental Microbiology, 2020, 22(7): 2469-2484.
doi: 10.1111/1462-2920.14961 |
| [3] | OLAITAN A O, MORAND S, ROLAIN J M. Mechanisms of polymyxin resistance: acquired and intrinsic resistance in bacteria. Frontier in Microbiology, 2014, 5: 643. |
| [4] | 宗劲, 徐凯进, 黄莹, 刘超群, 吴淼星, 韩东. 质粒介导黏菌素耐药机制研究进展. 中国微生态学杂志, 2021, 33(01): 120-125. |
| ZONG J, XU K J, HUANG Y, LIU C Q, WU M X, HAN D. Progress in research on plasmid-mediated colistin resistance. Chinese Journal of Microecology, 2021, 33(01): 120-125. (in Chinese) | |
| [5] |
LIU Y Y, WANG Y, WALSH T R, YI L X, ZHANG R, SPENCER J, DOI Y, TIAN G B, DONG B L, HUANG X H, YU L F, GU D X, REN H W, CHEN X, J LV L C, HE D D, ZHOU H W, LIANG Z S, LIU J H, SHEN J Z. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: a microbiological and molecular biological study. The Lancet Infectious Diseases, 2016, 16(2): 161-168.
doi: 10.1016/S1473-3099(15)00424-7 |
| [6] |
SUN J, ZHANG H M, LIU Y H, FENG Y J. Towards Understanding MCR-like Colistin Resistance. Trends in Microbiology, 2018, 26(9): 794-808.
doi: 10.1016/j.tim.2018.02.006 |
| [7] |
WANG R B, VAN D L, SHAW L P, BRADLEY P, WANG Q, WANG X J, JIN L Y, ZHANG Q, LIU Y Q, RIEUX A, DORAI-SCHNEIDERS T, WEINERT L A, IQBAL Z, DIDELOT X, WANG H, BALLOUX F. The global distribution and spread of the mobilized colistin resistance gene mcr-1. Nature Communications, 2018, 9(1): 1179.
doi: 10.1038/s41467-018-03205-z |
| [8] |
WANG Y, XU C Y, ZHANG R, CHEN Y Q, SHEN Y B, HU F P, LIU D J, LU J Y, GUO Y, XIA X, JIANG J Y, WANG X Y, FU Y L, YANG L, WANG J Y, LI J, CAI C, YIN D D, CHE J, FAN R, WANG Y Q, QING Y, LI Y, LIAO K, CHEN H, ZOU M X, LIANG L, TANG J, SHEN Z Q, WANG S L, YANG X R, WU C M, XU S X, WALSH T R, SHEN J Z. Changes in colistin resistance and mcr-1 abundance in Escherichia coli of animal and human origins following the ban of colistin-positive additives in China: an epidemiological comparative study. The Lancet Infectious Diseases, 2020, 20(10): 1161-1171.
doi: 10.1016/S1473-3099(20)30149-3 |
| [9] |
ANDERSSON D I, HUGHES D. Microbiological effects of sublethal levels of antibiotics. Nature Review Microbiology, 2014, 12(7): 465-478.
doi: 10.1038/nrmicro3270 |
| [10] |
LUNDSTROM S V, OSTMAN M, BENGTSSON-PALME J, RUTGERSSON C, THOUDAL M, SIRCAR T, BLANCK H, ERIKSSON K M, TYSKLIND M, FLACH C F, LARSSON D G J. Minimal selective concentrations of tetracycline in complex aquatic bacterial biofilms. Science of the Total Environment, 2016, 553: 587-595.
doi: 10.1016/j.scitotenv.2016.02.103 |
| [11] |
MARKOU N, MARKANTONIS S L, DIMITRAKIS E, PANIDIS D, BOUTZOUKA E, KARATZAS S, RAFAILIDIS P, APOSTOLAKOS H, BALTOPOULOS G. Colistin serum concentrations after intravenous administration in critically ill patients with serious multidrug-resistant, gram-negative bacilli infections: a prospective, open-label, uncontrolled study. Clinical Therapeutics, 2008, 30(1): 143-151.
doi: 10.1016/j.clinthera.2008.01.015 |
| [12] |
MOHAMED A F, KARAISKOS I, PLACHOURAS D, KARVANEN M, PONTIKIS K, JANSSON B, PAPADOMICHELAKIS E, ANTONIADOU A, GIAMARELLOU H, ARMAGANIDIS A, CARS O, FRIBERG L E. Application of a loading dose of colistin methanesulfonate in critically ill patients: population pharmacokinetics, protein binding, and prediction of bacterial kill. Antimicrobial Agents and Chemotherapy, 2012, 56(8): 4241-4249.
doi: 10.1128/AAC.06426-11 |
| [13] | The European Committee on Antimicrobial Susceptibility Testing. Breakpoint tables for interpretation of MICs and zone diameters, Version 11.0. http://www.eucast.org. 2021. |
| [14] | BETHKE J H, DAVIDOVICH A, CHENG L, LOPATKIN A J, SONG W, THADEN J T, FOWLER V G, J., XIAO M F, YOU L C. Environmental and genetic determinants of plasmid mobility in pathogenic Escherichia coli. Science Advances, 2020, 6(4): eaax3173. |
| [15] |
WAN Z, VARSHAVSKY J, TEEGALA S, MCLAWRENCE J, GODDARD NL. Measuring the rate of conjugal plasmid transfer in a bacterial population using quantitative PCR. Biophysical Journal, 2011, 101(1): 237-244.
doi: 10.1016/j.bpj.2011.04.054 |
| [16] | FU Y L, LIU D J, SONG H W, LIU Z H, JIANG H Y, WANG Y. Development of a Multiplex Real-Time PCR Assay for Rapid Detection of Tigecycline Resistance Gene tet(X) Variants from Bacterial, Fecal, and Environmental Samples. Antimicrobial Agents and Chemotherapy, 2020, 64(4): e02292-19. |
| [17] | LI J Y, SHI X M, YIN W J, WANG Y, SHEN Z Q, DING S Y, WANG S L. A Multiplex SYBR Green Real-Time PCR Assay for the Detection of Three Colistin Resistance Genes from Cultured Bacteria, Feces, and Environment Samples. Frontier in Microbiology, 2017, 8: 2078. |
| [18] | CHEN S F, ZHOU Y Q, CHEN Y R, GU J. fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics, 2018, 34(17): i884-i890. |
| [19] |
TJADEN B. A computational system for identifying operons based on RNA-seq data. Methods, 2020, 176: 62-70.
doi: 10.1016/j.ymeth.2019.03.026 |
| [20] |
KOPYLOVA E, NOE L, TOUZET H. SortMeRNA: fast and accurate filtering of ribosomal RNAs in metatranscriptomic data. Bioinformatics, 2012, 28(24): 3211-7.
doi: 10.1093/bioinformatics/bts611 |
| [21] |
KIM D, LANGMEAD B, SALZBERG S L. HISAT: a fast spliced aligner with low memory requirements. Nature Methods, 2015, 12(4): 357-360.
doi: 10.1038/nmeth.3317 |
| [22] |
LIAO Y, SMYTH G K, SHI W. featureCounts: an efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics, 2014, 30(7): 923-930.
doi: 10.1093/bioinformatics/btt656 |
| [23] |
LOVE M I, HUBER W, ANDERS S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biology, 2014, 15(12): 550.
doi: 10.1186/s13059-014-0550-8 |
| [24] | CABEZON E, ROZADA R J, PENA A, DE LA CRUZ F, ARECHAGA I. Towards an integrated model of bacterial conjugation. FEMS Microbiology Reviews, 2015, 39(1): 81-95. |
| [25] | RENDON M A, SALDANA Z, ERDEM A L, MONTEIRO-NETO V, VAZQUEZ A, KAPER J B, PUENTE J L, GIRON J A. Commensal and pathogenic Escherichia coli use a common pilus adherence factor for epithelial cell colonization. Proceedings of the National Academy of Sciences (USA), 2007, 104(25): 10637-10642. |
| [26] |
THOMAS C M, NIELSEN K M. Mechanisms of, and barriers to, horizontal gene transfer between bacteria. Nature Review Microbiology, 2005, 3(9): 711-721.
doi: 10.1038/nrmicro1234 |
| [27] |
GUO M T, YUAN Q B, YANG J. Distinguishing effects of ultraviolet exposure and chlorination on the horizontal transfer of antibiotic resistance genes in municipal wastewater. Environmental Science Technology, 2015, 49(9): 5771-5778.
doi: 10.1021/acs.est.5b00644 |
| [28] | QIU Z G, YU Y M, CHEN Z L, JIN M, YANG D, ZHAO Z G, WANG J, F SHEN Z Q, WANG X W, QIAN D, HUANG A H, ZHANG B C, LI J W. Nanoalumina promotes the horizontal transfer of multiresistance genes mediated by plasmids across genera. Proceedings of the National Academy of Sciences (USA), 2012, 109(13): 4944-4949. |
| [29] |
BEABER J W, HOCHHUT B, WALDOR M K. SOS response promotes horizontal dissemination of antibiotic resistance genes. Nature, 2004, 427(6969): 72-74.
doi: 10.1038/nature02241 |
| [30] |
JUTKINA J, RUTGERSSON C, FLACH C F, JOAKIM L D G. An assay for determining minimal concentrations of antibiotics that drive horizontal transfer of resistance. Science of the Total Environment, 2016, 548-549: 131-138.
doi: 10.1016/j.scitotenv.2016.01.044 |
| [31] |
KOCH-WESER J, SIDEL V W, FEDERMAN E B, KANAREK P, FINER D C, EATON A E. Adverse effects of sodium colistimethate. Manifestations and specific reaction rates during 317 courses of therapy. Annals of Internal Medicine, 1970, 72(6): 857-868.
doi: 10.7326/0003-4819-72-6-857 |
| [32] | AVALOS V I, HOSSEINI V, KOLLMANNSBERGER P, MEIER S, WEBER S S, ARNOLDINI M, ACKERMANN M, VOGEL V. How type1 fimbriae help Escherichia coli to evade extracellular antibiotics. Scientific Reports, 2016, 6: 18109. |
| [1] | ,,,. HPLC Analysis on Formation of the Multi-Bioinsecticidal-Toxin BtA [J]. Scientia Agricultura Sinica, 2005, 38(11): 2246-2253 . |
|
||